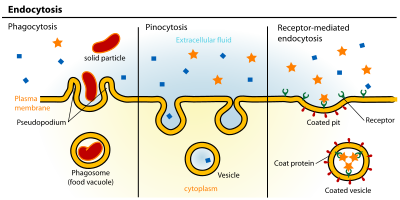
אנדוציטוזיס הוא תהליך סלולרי בו חומרים מובאים לתא. החומר שיש להפנים מוקף באזור של קרום תא, שאז נובט בתוך התא ויוצר a שַׁלפּוּחִית המכיל את החומר שנבלע. אנדוציטוזיס כולל פינוציטוזיס (שתיית תאים) ו פגוציטוזיס (אכילת תאים). זהו סוג של תחבורה פעילה.
הִיסטוֹרִיָה
המונח הוצע על ידי דה דוב בשנת 1963.[1] פגוציטוזיס התגלה על ידי אלי מטצ'ניקוף בשנת 1882.[2]
מסלולי אנדוציטוזה
ניתן לחלק את מסלולי האנדוציטוזה לארבע קטגוריות: אנדוציטוזה בתיווך קולטן (ידוע גם בשם אנדוציטוזה בתיווך קלתרין), caveolae, פינוציטוזיס, ו פגוציטוזיס.[3]
- אנדוציטוזה בתיווך קלתרין מתווכת על ידי ייצור שלפוחיות קטנות (כ- 100 ננומטר בקוטר) בעלות מעטה אופייני מורפולוגי המורכב מחלבון ציטוזולי קלתרין.[4] שלפוחיות מצופות קלתרין (CCVs) נמצאות כמעט בכל התאים ויוצרות תחומים של קרום הפלזמה המכונה בורות מצופים clathrin. בורות מצופים יכולים לרכז מולקולות חוץ-תאיות גדולות בעלות שונות קולטנים אחראי על האנדוציטוזה בתיווך קולטן של ליגנדים, למשל ליפופרוטאין בצפיפות נמוכה, טרנספרין, גורמי גדילה, נוגדנים ורבים אחרים.[5]
- לימוד [6] בתאי יונקים מאשרים הפחתה בגודל המעיל clathrin בסביבת מתח מוגברת. בנוסף, זה מציע ששני מצבי ההרכבה של קלתרין נבדלים, כלומר בורות מצופים ורובדים מצופים, שנצפו בחקירות ניסיוניות עשויים להיות תוצאה של מתחים מגוונים בקרום הפלזמה.
- Caveolae הם הנפוצים ביותר המדווחים על ניצני קרום הפלזמה שאינם מצופים קלתרין, אשר קיימים על פני השטח של רבים, אך לא כל סוגי התאים. הם מורכבים מחלבון המחייב כולסטרול קוואולין (Vip21) עם דו שכבתי מועשר ב כולסטרול ו גליקוליפידים. Caveolae הם קטנים (בקוטר של כ- 50 ננומטר) בורות בצורת בקבוקון בקרום הדומים לצורת מערה (ומכאן השם Caveolae). הם יכולים להוות עד שליש משטח קרום הפלזמה של התאים של רקמות מסוימות, בהיותם שופעים במיוחד שריר חלק, סוג I פנאומוציטים, פיברובלסטים, אדיפוציטים, ו תאי אנדותל.[7] האמונה כי ספיגת מולקולות חוץ-תאיות מתווכת באופן ספציפי באמצעות קולטנים במערות.
 מ- L ל- R: פגוציטוזיס, פינוציטוזיס, אנדוציטוזה בתיווך קולטנים.
מ- L ל- R: פגוציטוזיס, פינוציטוזיס, אנדוציטוזה בתיווך קולטנים.- פוטוציטוזיס היא צורה של אנדוציטוזה בתיווך קולטן המשתמשת בשלפוחיות caveolae בכדי להכניס מולקולות בגדלים שונים לתא. בניגוד לרוב האנדוציטוזה המשתמשת בקבולות למסירת תוכן של שלפוחית לליזוזומים או לאברונים אחרים, חומר המשולב באנדוציטוזיס באמצעות פוטוציטוזיס משתחרר לציטוזול.[8]
- פינוציטוזיס, המתרחש בדרך כלל מאזורים פרועים מאוד של קרום הפלזמה, הוא התפרצות קרום התא ליצירת כיס, ואז צובט בתא ליצירת שלפוחית (בקוטר של 5–5 מיקרומטר) מלא בנפח גדול של נוזל חוץ-תאי ומולקולות בתוכו (שווה ערך ל- ~ 100 CCV). מילוי הכיס מתרחש בצורה לא ספציפית. השלפוחית עוברת ואז אל תוך ציטוזול ונתיכים עם שלפוחיות אחרות כגון אנדוזומים ו ליזוזומים.[9]

- פגוציטוזיס הוא התהליך בו תאים נקשרים ומפנימים חומר חלקיקי שקוטרו גדול יותר מ- 0.75 מיקרומטר, כגון חלקיקי אבק קטנים, פסולת תאים, מיקרואורגניזמים ו אפופטוטי תאים. תהליכים אלה כרוכים בקליטת אזורי קרום גדולים יותר מאשר אנדוציטוזה בתיווך קלתרין ו caveolae נָתִיב.
ניסויים אחרונים יותר העלו כי תיאורים מורפולוגיים אלה של אירועים אנדוציטיים עשויים להיות בלתי מספקים, ושיטת סיווג מתאימה יותר עשויה להתבסס על תלות clathrin של מסלולים מסוימים, עם תת סוגים מרובים של endocytosis תלוי clathrin ו עצמאי clathrin. תובנה מכנית לאנדוציטוזה שאינה פגוציטית ואינה תלויה בקטרין הייתה חסרה, אך מחקר שנערך לאחרונה הראה כיצד גרף 1 מווסת נפוץ מאוד מסלול אנדוציטי בלתי תלוי בקתרין המכונה מסלול CLIC / GEEC.[10]
המרכיבים העיקריים של המסלול האנדוציטי
המסלול האנדוציטי של תאי יונקים מורכב מתאי קרום מובחנים, המפנימים מולקולות מקרום הפלזמה ומחזרים אותם חזרה אל פני השטח (כמו באנדוזומים מוקדמים ובמיחזור אנדוזומים), או ממיינים אותם עד השפלה (כמו באנדוזומים ומאוחרים בליזוזומים). המרכיבים העיקריים של המסלול האנדוציטי הם:[3]
- מוקדם אנדוזומים הם התא הראשון של המסלול האנדוציטי. אנדוזומים מוקדמים ממוקמים לעתים קרובות בפריפריה של התא, ומקבלים את רוב סוגי השלפוחיות המגיעות משטח התא. יש להם מבנה צינור-שלפוחית אופייני (שלפוחיות בקוטר של עד 1 מיקרומטר עם צינורות מחוברים בקוטר 50 ננומטר) ו- pH חומצי קל. הם בעיקר מיונים אברונים שבהם ליגנדים אנדוציטוזיים רבים מתנתקים מהם קולטנים ב- pH החומצי של התא, וממנו רבים מהקולטנים ממחזרים אל פני שטח התא (דרך צינורות).[11][12] זהו גם אתר המיון למסלול טרנסיטוטי לתאים מאוחרים יותר (כמו אנדוזומים מאוחרים או ליזוזומים) באמצעות תאים טרנסווסקולריים (כמו גופים רב-סיבוליים (MVB) או שלפוחיות נשא אנדוזומליות (ECV)).
- אנדוזומים מאוחרים לקבל חומר אנדוציטוז בדרך אל ליזוזומים, בדרך כלל מאנדוזומים מוקדמים במסלול האנדוציטי, מרשת טרנס-גולגי (TGN) במסלול הביוסינתטי, ומ פגוזומים במסלול הפאגוציטי.[13] אנדוזומים מאוחרים מכילים לעתים קרובות חלבונים האופייניים לנוקלאוזומים, מיטוכונדריה ו- mRNA כולל גליקופרוטאינים של קרום הליזוזומלי והידרולזים חומציים. הם חומציים (בערך pH 5.5), והם חלק ממסלול הסחר של מנוז -6 פוספט קולטנים. אנדוזומים מאוחרים נחשבים לתווך קבוצה אחרונה של אירועי מיון לפני העברת החומר לליזוזומים.
- ליזוזומים הם התא האחרון של המסלול האנדוציטי. תפקידם העיקרי הוא לפרק מוצרי פסולת תאית, שומנים, פחמימות, חלבונים ומקרומולקולות אחרות לתרכובות פשוטות. אלה מוחזרים לאחר מכן לציטופלסמה כחומרים חדשים לבניית תאים. כדי להשיג זאת, הליזוזומים משתמשים בכ- 40 סוגים שונים של אנזימים הידרוליטיים, שכולם מיוצרים ברטיקולום האנדופלזמי, שהשתנה מערכת גולג'י ולתפקד בסביבה חומצית.[14] ה- pH המשוער של ליזוזום הוא 4.8 ואילך אלקטרון מיקרוסקופי (EM) בדרך כלל נראים גדולים vacuoles (בקוטר 1-2 מיקרומטר) המכיל חומר צפוף אלקטרונים. יש להם תכולה גבוהה של חלבוני קרום ליזוזומליים והידרולזים ליזוזומליים פעילים, אך ללא קולטן מנוז -6 פוספט. הם נחשבים בדרך כלל לתא ההידרוליטי העיקרי של התא.[15][16]
לאחרונה נמצא כי eisosome משמש כפורטל לאנדוציטוזיס בשמרים.[17]
אנדוציטוזה בתיווך קלתרין
הדרך העיקרית לאנדוציטוזה ברוב התאים, והמובנת ביותר, היא המתווכת על ידי המולקולה קלתרין.[18][19] חלבון גדול זה מסייע ביצירת בור מצופה על פני השטח הפנימיים של קרום פלזמה של התא. לאחר מכן בור זה ניצב לתא ויוצר שלפוחית מצופה בציטופלזמה של התא. בכך הוא מכניס לתא לא רק שטח קטן של פני השטח של התא, אלא גם נפח קטן של נוזלים מחוץ לתא.[20][21][22]
מעילים מתפקדים כדי לעוות את קרום התורם לייצור שלפוחית, והם מתפקדים גם בבחירת מטען השלפוחית. מתחמי המעיל שאופיינו היטב עד כה כוללים חלבון מעיל I (COP-I), COP-II וקלתרין.[23][24] מעילי Clathrin מעורבים בשני שלבי תחבורה מכריעים: (i) אנדוציטוזה בתיווך קולטן ונוזל בשלב ממברנת הפלזמה לאנדוזום מוקדם ו- (ii) הובלה מה- TGN לאנדוזומים. באנדוציטוזיס, המעיל קלתרין מורכב על פני הציטופלזמה של קרום הפלזמה, ויוצר בורות שמתפשטים כדי לצבוט (נקרע) ולהפוך למשטחי CCV חופשיים. בתאים מתורבתים ההרכבה של CCV אורכת ~ דקה, וכמה מאות עד אלף ומעלה יכולים להיווצר בכל דקה.[25] מרכיב הפיגום העיקרי במעיל קלתרין הוא חלבון 190 קילוגרם הנקרא שרשרת כבדה קלתרין (CHC), אשר נקשר לחלבון 25 קילוגרם הנקרא שרשרת קלה קלתרין (CLC), ויוצר טרימרים בעלי שלוש רגליים הנקראים טריסקליונים.
שלפוחית להתרכז באופן סלקטיבי ולהוציא חלבונים מסוימים במהלך היווצרותם ואינם מייצגים את הממברנה כולה. מתאמי AP2 הם קומפלקסים רב-יחידתיים המבצעים פונקציה זו בקרום הפלזמה. הקולטנים המובנים ביותר שנמצאים מרוכזים בשלפוחיות מצופות של תאי יונקים הם קולטן LDL (שמסיר LDL מקולקולציה של דם), קולטן הטרנספרין (המביא ליוני ברזל קשורים טרנספרין לתוך התא) וקולטני הורמונים מסוימים (כמו למשל עבור EGF ).
בכל רגע, כ- 25% מקרום הפלזמה של פיברובלסט מורכב מבורות מצופים. מאחר ובור מצופה אורך חיים של כדקה לפני שהוא ניגן לתא, פיברובלסט לוקח את פניו בדרך זו בערך אחת ל -16 דקות. שלפוחיות מצופות הנוצרות מקרום הפלזמה יש קוטר של כ 36 ננומטר ואורך חיים נמדד בכמה שניות. לאחר שהמעיל נשפך, השלפוחית הנותרת מתמזגת עם אנדוזומים וממשיך במסלול האנדוציטי. תהליך הנביטה בפועל, לפיו בור הופך לשלפוחית, מתבצע על ידי קלתרין בסיוע מערכת חלבונים ציטופלזמית, הכוללת דינמין ומתאמים כגון אדפטין.
בורות וצפיפות מצופים נראו לראשונה בקטעים דקיקים של רקמות במיקרוסקופ האלקטרונים על ידי מאט אריות ופרקר ג'ורג '. את חשיבותם לפינוי LDL מדם גילה ריצ'רד ג 'אנדרסון, מייקל ס. בראון ו ג'וזף ל 'גולדשטיין בשנת 1977.[26] שלפוחית מצופה טוהרו לראשונה על ידי ברברה פירס, שגילה את מולקולת המעיל clathrin בשנת 1976.[27]
ראה גם
הפניות
- ^ ריגר, ר .; מיכאליס, א. גרין, מ.מ. 1991. מילון מונחים של גנטיקה. קלאסי ומולקולרי (מהדורה חמישית). ספרינגר-ורלאג, ברלין, [1].
- ^ "איליה מכניקוב - ביוגרפי". www.nobelprize.org. הוחזר 2016-10-10.
- ^ א ב מארש, מארק (2001). אנדוציטוזיס. הוצאת אוניברסיטת אוקספורד. עמ ' vii. ISBN 978-0-19-963851-2.
- ^ [2], McMahon, H. T. & Boucrot, E. מנגנון מולקולרי ותפקודים פיזיולוגיים של אנדוציטוזה בתיווך clathrin. סקירות טבע ביולוגיה של תאים מולקולריים 12, 517 (2011).
- ^ מארש, מ '; מקמהון, HT (יולי 1999). "העידן המבני של אנדוציטוזיס". מַדָע. 285 (5425): 215–20. דוי:10.1126 / science.285.5425.215. PMID 10398591.
- ^ אירג'יזאד, ה '; Agrawal, A. (2017). "פילמור של קלתרין מגלה רגישות מכאנית-גיאומטרית גבוהה". חומר רך. 13 (7): 1455–1462. דוי:10.1039 / C6SM02623K. PMC 5452080. PMID 28124714.
- ^ פרטון RG, סימונס K (מרץ 2007). "הפרצופים המרובים של הקבולות". סקירת טבע ביולוגיה של תאים מולקולריים. 8 (3): 185–94. דוי:10.1038 / nrm2122. PMID 17318224. S2CID 10830810.
- ^ מינאו, צ'יקו; אנדרסון, ריצ'רד ג'י (2001). "פוטוציטוזיס". היסטוכימיה וביולוגיה של תאים. 116 (2): 109–118. דוי:10.1007 / s004180100289. PMID 11685539.
- ^ Falcone S, Cocucci E, Podini P, Kirchhausen T, Clementi E, Meldolesi J (נובמבר 2006). "מקרופינוציטוזיס: תיאום מוסדר של אירועי תנועה בקרום אנדוציטיים ואקסוציטיים" (PDF). כתב העת למדעי התא. 119 (נק '22): 4758–69. דוי:10.1242 / jcs.03238. PMID 17077125. S2CID 14303429.
- ^ Lundmark R, Doherty GJ, Howes MT, et al. (נובמבר 2008). "החלבון GRAF1 המפעיל GTPF מסדיר את המסלול האנדוציטי CLIC / GEEC". ביולוגיה נוכחית. 18 (22): 1802–8. דוי:10.1016 / j.cub.2008.10.044. PMC 2726289. PMID 19036340.
- ^ מלמן הראשון (1996). "אנדוציטוזיס ומיון מולקולרי". סקירה שנתית של ביולוגיה של תאים והתפתחות. 12: 575–625. דוי:10.1146 / annurev.cellbio.12.1.575. PMID 8970738.
- ^ Mukherjee S, Ghosh RN, Maxfield FR (יולי 1997). "אנדוציטוזיס". ביקורות פיזיולוגיות. 77 (3): 759–803. דוי:10.1152 / physrev.1997.77.3.759. PMID 9234965.
- ^ Stoorvogel W, Strous GJ, Geuze HJ, Oorschot V, Schwartz AL (מאי 1991). "אנדוזומים מאוחרים נובעים מאנדוזומים מוקדמים על ידי התבגרות". תָא. 65 (3): 417–27. דוי:10.1016 / 0092-8674 (91) 90459-ג. PMID 1850321. S2CID 31539542.
- ^ ויסמן, ג '- מחקרים על ליזוזומים, 1965)
- ^ Gruenberg J, Maxfield FR (אוגוסט 1995). "הובלת ממברנה במסלול האנדוציטי". חוות דעת נוכחית בביולוגיה של תאים. 7 (4): 552–63. דוי:10.1016/0955-0674(95)80013-1. PMID 7495576.
- ^ לוזיו JP, Rous BA, Bright NA, PRryor PR, Mullock BM, Piper RC (1 במאי 2000). "היתוך ליסוזום-אנדוזום וביוגנזה של ליזוזום". כתב העת למדעי התא. 113 (9): 1515–24. PMID 10751143. הוחזר 2009-06-19.
- ^ Walther TC, Brickner JH, Aguilar PS, Bernales S, Pantoja C, Walter P (פברואר 2006). "Eisosomes מסמנים אתרים סטטיים של אנדוציטוזיס". טֶבַע. 439 (7079): 998–1003. דוי:10.1038 / nature04472. PMID 16496001. S2CID 2838121.
- ^ קירכהאוזן, ט.; אוון, ד '; הריסון, ס 'סי (1 במאי 2014). "מבנה מולקולרי, פונקציה ודינמיקה של תנועת ממברנה בתיווך קלתרין". פרספקטיבות נמל קרד ספרינג בביולוגיה. 6 (5): a016725. דוי:10.1101 / cshperspect.a016725. PMC 3996469. PMID 24789820.
- ^ ביציקאס, וי; Corrêa IR, ג'וניור; ניקולס, ביג '(17 בספטמבר 2014). "מסלולים שאינם תלויים בקלאתרין אינם תורמים באופן משמעותי לשטף האנדוציטי". eLife. 3: e03970. דוי:10.7554 / eLife.03970. PMC 4185422. PMID 25232658.
- ^ Benmerah A, Lamaze C (אוגוסט 2007). "בורות מצופים קלתרין: vive la différence?". תְנוּעָה. 8 (8): 970–82. דוי:10.1111 / j.1600-0854.2007.00585.x. PMID 17547704.
- ^ Rappoport JZ (יוני 2008). "התמקדות באנדוציטוזה בתיווך קלתרין". כתב העת הביוכימי. 412 (3): 415–23. דוי:10.1042 / BJ20080474. PMID 18498251. S2CID 24174632.
- ^ גרנשת ב ', אודרמאט ב', רויל ס.ג ', לאגנדו ל' (דצמבר 2007). "אנדוציטוזה בתיווך קלתרין: המנגנון הפיזיולוגי של שליפת שלפוחית בסינפסות בהיפוקמפוס". כתב העת לפיזיולוגיה. 585 (נק '3): 681–6. דוי:10.1113 / jphysiol.2007.139022. PMC 2375507. PMID 17599959.
- ^ רובינסון טרשת נפוצה (מרץ 1997). "מעילים וצמיחת שלפוחית". מגמות בביולוגיה של תאים. 7 (3): 99–102. דוי:10.1016 / S0962-8924 (96) 10048-9. PMID 17708916.
- ^ גליק BS, Malhotra V (דצמבר 1998). "מעמדו המוזר של מנגנון גולגי". תָא. 95 (7): 883–9. דוי:10.1016 / S0092-8674 (00) 81713-4. PMID 9875843.
- ^ גאידרוב הראשון, סנטיני ו ', וורן ר.א., קין ג'ה (מאי 1999). "שליטה מרחבית על דינמיקת בור מצופה בתאים חיים". ביולוגיה של תאי טבע. 1 (1): 1–7. דוי:10.1038/8971. PMID 10559856. S2CID 12553151.
- ^ אנדרסון, RG; בראון, טרשת נפוצה; גולדשטיין, JL (מרץ 1977). "תפקיד השלפוחית האנדוציטית המצופה בקליטת ליפופרוטאין בצפיפות נמוכה הקשורה לקולטן בפיברובלסטים אנושיים". תָא. 10 (3): 351–64. דוי:10.1016/0092-8674(77)90022-8. PMID 191195. S2CID 25657719.
- ^ Pearse, BM (אפריל 1976). "קלתרין: חלבון ייחודי הקשור להעברה קרומית של תאית על ידי שלפוחית מצופה". הליכים של האקדמיה הלאומית למדעים של ארצות הברית של אמריקה. 73 (4): 1255–9. דוי:10.1073 / pnas.73.4.1255. PMC 430241. PMID 1063406.