ב ביולוגיה מולקולרית, ה מכסה של חמש פריים (מכסה 5 ') הוא שינוי מיוחד נוקלאוטיד על סוף 5 ′ של חלקם תמלילים ראשוניים כמו מבשר RNA שליח. תהליך זה, המכונה מכסת mRNA, מאוד מוסדר וחיוני ביצירת יציבות ו RNA שליח בוגר מסוגל לעבור תִרגוּם בְּמַהֲלָך סינתזת חלבונים. מיטוכונדריאלי mRNA[1] ו כלורופלסטיקה mRNA[2] אינם מוגבלים.
מִבְנֶה

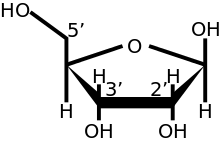
ב אקריוטים, מכסה 5 '(cap-0), שנמצא בקצה 5' של מולקולת mRNA, מורכב גואנין נוקלאוטיד המחובר ל- mRNA באמצעות 5 'עד 5' יוצא דופן טריפוספט הַצמָדָה. זֶה גואנוזין הוא מתיל בעמדה 7 מיד לאחר המכסה in vivo על ידי א מתילטרנספרז.[3][4][5][6] זה מכונה א 7-מתילגואנילט מכסה, מקוצר m7ז.
באיקריוטים רב-תאיים ובנגיפים מסוימים,[7] קיימים שינויים נוספים, כולל מתילציה של ה -2 ' קבוצות הידרוקסי של 2 הראשונים ריבוז סוכרים בקצה 5 ′ של ה- mRNA. ל- cap-1 יש קבוצת 2'-הידרוקסי מתילית על סוכר הריבוז הראשון, ואילו ל- cap-2 קבוצות של 2'-הידרוקסיות מתיליות בשני הסוכרים הראשונים של הריבוז, המוצגים מימין. מכסה 5 ′ דומה מבחינה כימית ל סוף 3 ′ של מולקולת RNA (פחמן 5 ′ של הכובע ריבוז נקשר, וה 3 ′ לא קשור). זה מספק עמידות משמעותית ל 5 ' exonucleases.[צורך בציטוט ]
RNAים גרעיניים קטנים מכילים 5 'כובעים ייחודיים. סוג SNRNA של סוג SM נמצא עם כובעי 5'-trimethylguanosine, ואילו snRNAs מסוג Lsm נמצאים עם מכסי 5'-monomethylethylphosphate.[8]
ב בַּקטֶרִיָה, ואולי גם באורגניזמים גבוהים יותר, כמה רנ"א מוגבלים נאד+, NADH, או 3'-דה-פוספו-קואנזים A.[9][10]
בכל האורגניזמים ניתן לפרוק את מולקולות ה- mRNA בתהליך המכונה מסיר RNA decapping.
תהליך מכסה
נקודת המוצא לכיסוי עם 7-מתיל-גואנילט היא קצה 5 'ללא שינוי של מולקולת RNA, אשר מסתיימת בקבוצת טריפוספט. זה כולל נוקלאוטיד סופי ואחריו שלוש קבוצות פוספט המחוברות לפחמן 5 ′.[3] תהליך המכסת מתחיל לפני השלמת התמלול, מכיוון שמסינתזת ה- pre-mRNA המתהווה.
- אחת מקבוצות הפוספט הסופיות מוסרת על ידי טריפוספטאז RNA, משאיר קבוצת ביספוספט (כלומר 5 ′ (ppN) [pN]נ);
- GTP מתווסף לביספוספט הסופי על ידי mRNA guanylyltransferase, לאבד א פירופוספט ממצע ה- GTP בתהליך. התוצאה היא הצמדה של טריפוספט 5'– 5 ', המייצרת 5' (Gp) (ppN) [pN]נ;
- 7-חנקן של גואנין הוא מתיל על ידי mRNA (גואנין-נ7 -) - מתילטרנספרז, עם ס-adenosyl-ל-מתיונין להיות demethylated לייצר ס-adenosyl-ל-הומוציסטאין, וכתוצאה מכך 5 '(m7Gp) (ppN) [pN]נ (כובע -0);
- שינויים סמוכים למכסה יכולים להתרחש, בדרך כלל לנוקליאוטידים הראשונים והשניים, ומייצרים עד 5 '(m7Gp) (ppN *) (pN *) [pN]נ (כובע -1 וכובע -2);[7]
- אם הנוקלאוטיד הסמוך לכובע הוא 2′-או-ריבוז מתיל אדנוזין (כלומר 5 '(m7Gp) (ppAm) [pN]נ), זה יכול להיות מתיל נוסף במצב מתיל N6 כדי ליצור נ6-מתילאדנוזין, וכתוצאה מכך 5 '(m7Gp) (ppm6Am) [pN]נ.[3]
מנגנון המכסת עם NAD+, NADH או 3'-dephospho-coenzyme A שונה. מכסת עם NAD+, NADH או 3'-dephospho-coenzyme A מושג באמצעות "מנגנון מכסת ab initio", שבו NAD+, NADH או 3'-desphospho-coenzyme A משמש כ"נוקליאוטיד יוזם לא קנוני "(NCIN) עבור חניכת תמלול על ידי RNA פולימראז ועל ידי כך הוא משולב ישירות במוצר ה- RNA.[9] גם פולימראז RNA חיידקי וגם אוקריוטי RNA פולימראז II מסוגלים לבצע את "מנגנון המכסת ab initio" הזה.[9]
מיקוד
עבור מכסה עם 7-methylguanylate, אנזים מכסת מורכב (CEC) נקשר ל RNA פולימראז II לפני תחילת התמלול. ברגע שקצה 5 ′ של התמליל החדש יוצא מ- RNA פולימראז II, ה- CEC מבצע את תהליך המכסת (מנגנון מסוג זה מבטיח מכסת, כמו עם polyadenylation ).[11][12][13][14] האנזימים למכסה יכולים להיקשר רק ל RNA פולימראז II, להבטיח ספציפיות רק לתמלילים אלה, שהם כמעט לחלוטין mRNA.[12][14]
מכסת עם NAD+, NADH או 3'-dephospho-coenzyme A ממוקד על ידי מְקַדֵם סדר פעולות.[9] מכסת עם NAD +, NADH או 3'-dephospho-coenzyme A מתרחשת רק אצל מקדמים שיש להם רצפים מסוימים ומיד במעלה הזרם של אתר התחלת השעתוק ולכן מתרחשים רק עבור RNAs המסונתזים ממקדמים מסוימים.[9]
פוּנקצִיָה
מכסה 5 ′ יש ארבע פונקציות עיקריות:
- ויסות יצוא גרעיני;[15][16]
- מניעת השפלה על ידי exonucleases;[9][17][18][19]
- קידום תרגום (ראה ריבוזום ו תִרגוּם );[3][4][5]
- קידום כריתה אינטרונית פרוקסימאלית.[20]
ייצוא גרעיני של RNA מוסדר על ידי מתחם כריכת כובע (CBC), הנקשר באופן בלעדי ל- RNA מכוסה 7-מתיל-גואנילט. ה- CBC מוכר אז על ידי מתחם נקבוביות גרעיניות ומיוצא. לאחר הציטופלסמה לאחר סבב התרגום החלוצי, ה- CBC מוחלף בגורמי התרגום eIF4E ו eIF4G של ה eIF4F מורכב.[6] מתחם זה מוכר על ידי מכונות ייזום תרגום אחרות, כולל הריבוזום.[21]
מכסה עם 7-methylguanylate מונע השפלה של 5 'בשתי דרכים. ראשית, השפלה של ה- mRNA על ידי אקסונוקליזות 5 'מונעת (כאמור לעיל) על ידי מראה פונקציונלי כמו קצה 3'. שנית, ה- CBC ו- eIF4E / eIF4G חוסמים את הגישה של אנזימים הניתנים לניתוק מכסה. זה מגדיל את חצי חיים של ה- mRNA, החיוני באיקריוטים מכיוון שתהליכי הייצוא והתרגום אורכים זמן משמעותי.
Decapping של 7-methylguanylate מכוסה mRNA מזרז על ידי מתחם decapping מורכב לפחות Dcp1 ו Dcp2, אשר חייב להתחרות עם eIF4E כדי לאגד את המכסה. לפיכך מכסה 7-מתילגואנילט הוא סמן של mRNA המתורגם באופן פעיל ומשמש תאים לוויסות מחצית חיים של mRNA בתגובה לגירויים חדשים. MRNAs לא רצויים נשלחים אל גופי P לאחסון או decapping זמני, שפרטיהם עדיין נפתרים.[22]
המנגנון של קידום כריתה אינטרונית פרוקסימלית 5 'אינו מובן היטב, אך נראה כי מכסה 7-מתיל-גואנילט מסתובב ומתקשר עם אחוי בתהליך השבחה, קידום כריתה באינטרון.
ראה גם
הפניות
- ^ Temperley RJ, Wydro M, Lightowlers RN, Chrzanowska-Lightowlers ZM (יוני 2010). "MRNAs מיטוכונדריאליים אנושיים - כמו בני כל המשפחות, דומים אך שונים". Biochimica et Biophysica Acta (BBA) - ביואנרגטיקה. 1797 (6–7): 1081–1085. דוי:10.1016 / j.bbabio.2010.02.036. PMC 3003153. PMID 20211597.
- ^ מונה RA, שוסטר G, שטרן DB (7 ביוני 2000). "עיבוד והשפלה של mRNA כלורופלסט". ביוכימי. 82 (6–7): 573–582. דוי:10.1016 / S0300-9084 (00) 00606-4. PMID 10946108.
- ^ א ב ג ד שטקין, א '(דצמבר 1976). "מכסת mRNAs אוקריוטים". תָא. 9 (4): 645–653. דוי:10.1016/0092-8674(76)90128-8. PMID 1017010. S2CID 26743858.
- ^ א ב Banerjee AK (יוני 1980). "מבנה מכסה 5'-סופני בחומצות ריבונוקלאיות של שליח אוקריוטי". ביקורות מיקרוביולוגיות. 44 (2): 175–205. דוי:10.1128 / mmbr.44.2.175-205.1980. PMC 373176. PMID 6247631.
- ^ א ב Sonenberg N, Gingras AC (אפריל 1998). "ה- mRNA 5 'חלבון המחייב כובע eIF4E ושליטה בצמיחת תאים". חוות דעת נוכחית בביולוגיה של תאים. 10 (2): 268–275. דוי:10.1016 / S0955-0674 (98) 80150-6. PMID 9561852.
- ^ א ב Marcotrigiano J, Gingras AC, Sonenberg N, Burley SK (יוני 1997). "מבנה קוקיסטל של החלבון המחייב כובע של שליח RNA 5 '(eIF4E) קשור ל- 7-methyl-GDP". תָא. 89 (6): 951–961. דוי:10.1016 / S0092-8674 (00) 80280-9. PMID 9200613. S2CID 15200116.
- ^ א ב פכטר פ, בראונלי GG (מאי 2005). "הכרה במבני כובע mRNA על ידי חלבונים נגיפיים ותאים". כתב העת לווירולוגיה כללית. 86 (נק '5): 1239-1249. דוי:10.1099 / vir.0.80755-0. PMID 15831934. הועבר לארכיון מ המקורי בתאריך 07/06/2013. הוחזר 2014-12-12.
- ^ מטרה AG, טרנס RM, חבר הפרלמנט של טרנס (מרץ 2007). "רנ"א שאינם מקודדים: לקחים מרנ"א גרעיני קטן ומקטעי רנ"א קטנים". ביקורות טבע. ביולוגיה של תאים מולקולריים. 8 (3): 209–220. דוי:10.1038 / nrm2124. PMID 17318225. S2CID 30268055.
- ^ א ב ג ד ה f Bird JG, Zhang Y, Tian Y, Panova N, Barvík I, Greene L, Liu M, Buckley B, Krásný L, Lee JK, CD Kaplan, Ebright RH, Nickels BE (יולי 2016). "המנגנון של מכסת RNA 5 'עם NAD +, NADH ו- desphospho-CoA". טֶבַע. 535 (7612): 444–447. ביבקוד:2016Natur.535..444B. דוי:10.1038 / nature18622. PMC 4961592. PMID 27383794.
- ^ Cahová H, Winz ML, Höfer K, Nübel G, Jäschke A (מרץ 2015). "NAD captureSeq מציין NAD ככובע חיידקי עבור תת קבוצה של RNAs רגולטוריים". טֶבַע. 519 (7543): 374–377. ביבקוד:2015 טבע 519..374 ג. דוי:10.1038 / nature14020. PMID 25533955. S2CID 4446837.
- ^ צ'ו EJ, Takagi T, מור CR, Buratowski S (דצמבר 1997). "אנזים מכסת ה- mRNA מגויס למתחם התעתיק על ידי זרחון של תחום ה- RNA פולימראז II המסוף לקרבוקסי". גנים ופיתוח. 11 (24): 3319–3326. דוי:10.1101 / gad.11.24.3319. PMC 316800. PMID 9407025.
- ^ א ב Fabrega C, Shen V, Shuman S, CD של לימה (יוני 2003). "מבנה של אנזים מכסת mRNA הקשור לתחום המסוף של קרבוקסי בזרחן של RNA פולימראז II". תא מולקולרי. 11 (6): 1549–1561. דוי:10.1016 / S1097-2765 (03) 00187-4. PMID 12820968.
- ^ Ho CK, Lehman K, Shuman S (דצמבר 1999). "מוטיב משטח חיוני (WAQKW) של טריפוספטאז RNA שמרים מתווך היווצרות קומפלקס האנזים מכסת ה- mRNA עם RNA guanylyltransferase". מחקר על חומצות גרעין. 27 (24): 4671–4678. דוי:10.1093 / נאר / 27.24.4671. PMC 148765. PMID 10572165.
- ^ א ב Hirose Y, Manley JL (יוני 2000). "RNA פולימראז II ושילוב אירועים גרעיניים". גנים ופיתוח. 14 (12): 1415–1429. דוי:10.1101 / gad.14.12.1415 (לא פעיל 2020-09-01). PMID 10859161. הוחזר 23 בנובמבר 2014.CS1 maint: DOI לא פעיל החל מספטמבר 2020 (קישור)
- ^ ויזה נ ', איזאורלדה ה', פריירה ג'יי, דנהולט ב ', מטאג' IW (אפריל 1996). "קומפלקס מחייב כובע גרעיני קושר את טבעת הבלביאני לפני ה- mRNA באופן תעתיק ומלווה את חלקיק הריבונוקלאופרוטאין במהלך הייצוא הגרעיני.". כתב העת לביולוגיה של תאים. 133 (1): 5–14. דוי:10.1083 / jcb.133.1.5. PMC 2120770. PMID 8601613.
- ^ לואיס JD, איזאורלדה E (יולי 1997). "תפקידו של מבנה המכסה בעיבוד RNA וייצוא גרעיני". כתב העת האירופי לביוכימיה. 247 (2): 461–469. דוי:10.1111 / j.1432-1033.1997.00461.x. PMID 9266685.
- ^ Evdokimova V, Ruzanov P, Imataka H, Raught B, Svitkin Y, Ovchinnikov LP, Sonenberg N (אוקטובר 2001). "החלבון העיקרי הקשור ל- mRNA YB-1 הוא מייצב mRNA חזק התלוי בכובע 5 '". כתב העת EMBO. 20 (19): 5491–5502. דוי:10.1093 / emboj / 20.19.5491. PMC 125650. PMID 11574481.
- ^ Gao M, Fritz DT, Ford LP, Wilusz J (מרץ 2000). "אינטראקציה בין ריבונוקלאז פולי (A) ספציפי לבין מכסה 5 'משפיעה על שיעורי דדנילציה של mRNA במבחנה". תא מולקולרי. 5 (3): 479–488. דוי:10.1016 / S1097-2765 (00) 80442-6. PMC 2811581. PMID 10882133.
- ^ בורקארד KT, באטלר JS (ינואר 2000). "אקסונוקלאז גרעיני 3'– 5 'המעורב בפירוק ה- mRNA מתקשר עם פולימראז פולי (A) וחלבון ה- hnRNA Npl3p". ביולוגיה מולקולרית ותאית. 20 (2): 604–616. דוי:10.1128 / MCB.20.2.604-616.2000. PMC 85144. PMID 10611239.
- ^ Konarska MM, Padgett RA, Sharp PA (אוקטובר 1984). "הכרה במבנה הכובע בשחבור במבחנה של קודמי ה- mRNA". תָא. 38 (3): 731–736. דוי:10.1016 / 0092-8674 (84) 90268-X. PMID 6567484. S2CID 10721149.
- ^ Kapp LD, Lorsch JR (2004). "המכניקה המולקולרית של התרגום האוקריוטי". סקירה שנתית של ביוכימיה. 73 (1): 657–704. דוי:10.1146 / annurev.biochem.73.030403.080419. PMID 15189156.
- ^ פארקר R, שת U (מרץ 2007). "גופי P ושליטה בתרגום והשפלה של mRNA". תא מולקולרי. 25 (5): 635–646. דוי:10.1016 / j.molcel.2007.02.011. PMID 17349952.
קישורים חיצוניים
- "כובעי RNA". כותרת נושא רפואי של PubMed (MeSH). המכונים הלאומיים לבריאות.