| מכרסם | |
|---|---|
 | |
| עם כיוון השעון משמאל למעלה: קפיברה, ארבע קפיץ, סנאי קרקע עטוף זהב, עכבר הבית ו בונה בצפון אמריקה המייצג את תת-הסדרים Hystricomorpha, Anomaluromorpha, Sciuromorpha, Myomorpha ו- Castorimorpha בהתאמה. | |
| סיווג מדעי | |
| מַלְכוּת: | בעלי חיים |
| מַעֲרָכָה: | Chordata |
| מעמד: | ממליה |
| (לא מדורג): | גלירס |
| להזמין: | רודנטיה Bowdich, 1821 |
| סדרי משנה | |
אנומלורומורפה | |
 | |
| מגוון משולב של כל מיני המכרסמים (לא כולל אוכלוסיות שהוכנסו) | |
מכרסמים (מ לָטִינִית רודר, "לכרסם") הם יונקים של ה להזמין רודנטיה (/רoʊˈדɛנʃə/), המאופיינים בזוג יחיד של צמיחה מתמדת חותכות בכל אחד מהעליונים והתחתונים לסתות. כ -40% מכל מיני היונקים הם מכרסמים; הם נמצאים במספרים עצומים בכל היבשות למעט אנטארקטיקה. הם סדר היונקים המגוון ביותר וחיים במגוון ארציות בתי גידול, כולל סביבות מעשה ידי אדם.
מינים יכולים להיות arboreal, פוסוריאלי (נבירה), או חצי-מים. מכרסמים ידועים כוללים עכברים, חולדות, סנאים, כלבי ערבה, שבבי שבבים, צ'ינצ'ילות, דורבנים, בונים, שרקנים, אוגרים, גרבילים ו קפיברות. ארנבות, ארנבות, ו פיקאות, שגם השיניים החותכות שלהם צומחות ללא הרף, נכללו בהן פעם, אך נחשבים כעת לסדר נפרד, לגומורפה. עם זאת, רודנטיה ולאגומורפה כן קבוצות אחות, שיתוף א אב קדמון משותף יחיד ויוצרים את לבוש שֶׁל גלירס.
רוב המכרסמים הם בעלי חיים קטנים עם גופים חזקים, גפיים קצרות וזנבות ארוכים. הם משתמשים בשיניים החותכות שלהם כדי לכרסם מזון, לחפור מאורות ולהגן על עצמם. רובם אוכלים זרעים או חומר צמחי אחר, אך בחלקם יש דיאטות מגוונות יותר. הם נוטים להיות חיות חברתיות ומינים רבים חיים בחברות עם דרכים מורכבות לתקשר זה עם זה. ההזדווגות בין מכרסמים יכולה להשתנות בין מוֹנוֹגָמִיָה, ל פוליגניה, ל הֶפקֵרוּת. לרבים יש המלטות של לא מפותח, אלטריאלי צעירים, בעוד שאחרים הם טרום חברתי (מפותח יחסית) בלידה.
המכרסם מְאוּבָּן רשומה תאריכים חזרה פליאוקן ביבשת העל של לוראסיה. מכרסמים גוונו מאוד באזור אאוקן, כשהם התפשטו על פני יבשות, לפעמים אפילו חציית אוקיינוסים. מכרסמים הגיעו מדרום אמריקה ומדגסקר מאפריקה ועד בואם הומו סאפיינס, היו הארציים היחידים שליה יונקים להגיע אוסטרליה להתיישב בה.
מכרסמים שימשו כמזון, לבגדים, כחיות מחמד וכמו חיות מעבדה במחקר. מינים מסוימים, בפרט, עכברוש חום, ה חולדה שחורה, וה עכבר הבית, הם רציניים מזיקים, אכילה וקלקול מזון המאוחסן על ידי בני אדם והפצת מחלות. בטעות מינים שהוצגו של מכרסמים נחשבים לעתים קרובות להיות פַּלשָׁנִי וגרמו להכחדתם של מינים רבים, כגון ציפורי איים, שבודדו בעבר מטורפים יבשתיים.
מאפיינים
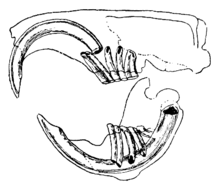
המאפיין המובהק של המכרסמים הוא זוגותיהם הצומחים באופן חד, חד כתער, שורשים פתוחים חותכות.[1] בשיניים החותכות הללו יש שכבות אמייל עבות מלפנים ומעט אמייל מאחור.[2] מכיוון שהם לא מפסיקים לגדול, על בעל החיים להמשיך לשחוק אותם כדי שלא יגיעו וינקבו את הגולגולת. כאשר השיניים החותכות טוחנות זו בזו, השיניים הרכות בחלק האחורי של השיניים נשחקות ומשאירות את קצה האמייל החד בצורת להב של אִזְמֵל.[3] לרוב המינים יש עד 22 שיניים ללא כלבים או קדמיים מקדים. פער, או דיאסטמה, מתרחשת בין השיניים החותכות לשיני הלחי ברוב המינים. זה מאפשר למכרסמים לינוק בלחיים או בשפתיים כדי להגן על פיהם וגרונם מפני שבבי עץ וחומר בלתי אכיל אחר, ומשליך פסולת זו מצידי פיהם.[4] לצ'ינצ'ילות וחזירי ים יש תזונה עשירה בסיבים; לטוחנות שלהם אין שורשים וגדלים ברציפות כמו החותכות שלהם.[5]
אצל מינים רבים, הטוחנות גדולות יחסית, מובנות מורכבות ומבולבלות או מחורצות מאוד. טוחנות מכרסמים מצוידות היטב לטחינת מזון לחלקיקים קטנים.[1] שרירי הלסת חזקים. הלסת התחתונה נדחקת קדימה בזמן מכרסם ונמשכת לאחור במהלך הלעיסה.[2] קבוצות מכרסמים נבדלות בסידור שרירי הלסת ומבני הגולגולת הנלווים, הן מיונקים אחרים והן בינם לבין עצמם. ה Sciuromorpha, כמו ה סנאי אפור מזרחי, יש עמוק גדול מטבח, מה שהופך אותם ליעילים בנשיכות עם החותכות. ה מיומורפה, כמו החולדה החומה, התרחבו temporalis השרירים, מה שמאפשר להם ללעוס בעוצמה בעזרת הטוחנות שלהם. ה היסטריקומורפה, כמו שפן הניסיונות, הם בעלי שרירי טריטור שטחיים גדולים יותר ושרירי טריטור עמוקים קטנים יותר מאשר חולדות או סנאים, ואולי הופכים אותם ליעילים פחות בכריכה עם החותכות, אך שרירי הפטריגואיד הפנימיים המוגדלים שלהם עשויים לאפשר להם להעביר את הלסת הצידה יותר בעת לעיסה. .[6] ה שקית לחיים הוא מאפיין מורפולוגי ספציפי המשמש לאחסון מזון והוא ניכר בתתי קבוצות מסוימות של מכרסמים כמו חולדות קנגורו, אוגרים, שבבי שבבים וגופרים שיש להם שתי שקיות שעשויות לנוע בין הפה לקדמת הכתפיים.[7] עכברים וחולדות אמיתיים אינם מכילים מבנה זה אך לחייהם אלסטיות בגלל רמה גבוהה של שרירים ועצבנות באזור.[8]

בעוד שהמין הגדול ביותר, קפיברה, יכול לשקול עד 66 ק"ג (146 ליברות), רוב המכרסמים שוקלים פחות מ 100 גרם (3.5 עוז). המכרסם הקטן ביותר הוא בלוכיסטאן פיגמי ג'רבואה הממוצע הוא 4.4 ס"מ בלבד (1.7 אינץ ') באורך הראש והגוף, כאשר הנקבות הבוגרות שוקלות 3.75 גרם בלבד (0.132 עוז). למכרסמים יש מורפולוגיות רחבות טווח, אך בדרך כלל יש גופות גוץ וגפיים קצרות.[1] בגפיים הקדמיות יש בדרך כלל חמש ספרות, כולל אגודל מנוגד, ואילו בגפיים האחוריות יש שלוש עד חמש ספרות. המרפק מעניק לאמות היד גמישות רבה.[3][9] רוב המינים הם צמחייה, הולכים על כפות הידיים ועל כפות הרגליים, ויש להם ציפורניים דומות לטופר. ציפורניים של מינים נוברים נוטות להיות ארוכות וחזקות, ואילו למכרסמים בעץ ציפורניים קצרות וחדות יותר.[9] מיני מכרסמים משתמשים במגוון רחב של שיטות תנועה כולל מרובע הליכה, ריצה, נבירה, טיפוס, דו-כיווני מקפץ (חולדות קנגורו ו עכברים מקפצים ), שחייה ואפילו גלישה.[3]סנאים קשקשים ו סנאים מעופפים, למרות שאינם קשורים זה לזה, שניהם יכולים להחליק מעץ לעץ באמצעות קרומים דמויי מצנח הנמתחים מקדימה לגפיים האחוריות.[10] ה אגוטי הוא רגל צי ו אַנְטִילוֹפָּה כמו, להיות digitigrade ובעלי ציפורניים דמויות פרסה. לרוב המכרסמים יש זנבות, שיכולים להיות בעלי צורות וגדלים רבים. כמה זנבות הם מתיחה מקדימה, כמו ב עכבר הקציר האירואסי, והפרווה על הזנבות יכולה להשתנות בין עבות לקרחת לחלוטין. הזנב משמש לעיתים לתקשורת, כמו כאשר ביברים מטיחים את זנבותיהם על פני המים או שעכברי בית מקשקש בזנבותיהם כדי להצביע על אזעקה. במינים מסוימים יש זנבות פרוזדוריים או ללא זנבות כלל.[1] אצל מינים מסוימים הזנב מסוגל להתחדש אם חלק נשבר.[3]

למכרסמים יש בדרך כלל חושים מפותחים רֵיחַ, שמיעה, וראייה. למינים ליליים יש לעתים קרובות עיניים מוגדלות וחלקם רגישים ל אוּלְטרָה סָגוֹל אוֹר. למינים רבים יש רגישים ארוכים שָׂפָם או ויברציות למגע או "מקציף". לחלק מהמכרסמים יש שקיות לחיים, שעשוי להיות מרופד בפרווה. אלה יכולים להיות מופנים כלפי חוץ לצורך ניקוי. אצל מינים רבים, הלשון לא יכולה להגיע אל עבר החותכות. למכרסמים מערכות עיכול יעילות, סופגות כמעט 80% מהאנרגיה הנבלעת. כשאוכלים תָאִית המזון מתרכך בבטן ומועבר ל שסתום, שם חיידקים מפחיתים אותו ל פַּחמֵימָה אלמנטים. המכרסם ואז מתרגל קופרופגיה, אוכלת כדורי צואה משלה, כך שהרכיבים התזונתיים יכולים להיספג במעיים. לכן מכרסמים מייצרים לעתים קרובות גלולה צואתית קשה ויבשה.[1] אצל מינים רבים, הפין מכיל עצם, baculum; האשכים יכולים להיות ממוקמים בבטן או במפשעה.[3]
דימורפיזם מיני מופיע אצל מיני מכרסמים רבים. בחלק מהמכרסמים הזכרים גדולים מנקבות ואילו אצל אחרים ההפך הוא הנכון. דימורפיזם מיני של הטיה גברית אופייני ל סנאים קרקעיים, חולדות קנגורו, בודדות חולדות שומה ו גופרים בכיס; זה כנראה התפתח בגלל סלקציה מינית ולחימה גברית-גברית גדולה יותר. דימורפיזם מיני של הטיה נשית קיים בקרב שבבי שבבים ו עכברים קופצים. לא מובן מדוע דפוס זה מתרחש, אך במקרה של שבבי צהוב-אורן, גברים עשויים לבחור נקבות גדולות יותר בשל הצלחתן הרבה יותר. במינים מסוימים, כגון מטחנים, דימורפיזם מיני יכול להשתנות מאוכלוסיה לאוכלוסייה. ב כספי בנק נקבות בדרך כלל גדולות יותר מזכרים, אך דימורפיזם מיני של הטיה גברית מתרחש באוכלוסיות אלפיניות, אולי בגלל היעדר טורפים ותחרות גדולה יותר בין גברים.[11]
הפצה ובית גידול

אחת הקבוצות הנפוצות ביותר של יונקים, מכרסמים ניתן למצוא בכל יבשת פרט לאנטארקטיקה. הם היונקים השיליים הארציים היחידים שהתיישבו באוסטרליה גינאה החדשה ללא התערבות אנושית. בני אדם אפשרו גם לבעלי החיים להתפשט לאיים אוקיאניים מרוחקים רבים (למשל חולדה פולינזית ).[3] מכרסמים הסתגלו כמעט לכל בית גידול ארצי, מקור טונדרה (שם הם יכולים לחיות מתחת לשלג) למדבריות לוהטות.
מינים מסוימים כגון סנאי עץ ו דורבנים של העולם החדש הם arboreal, בעוד שחלקם, כגון גופרים, טוקו-טוקו וחולדות שומות, חיות כמעט מתחת לאדמה, שם הן בונות מערכות מחילה מורכבות. אחרים שוכנים על פני האדמה, אך עשויים להיות להם מחילה אליה הם יכולים לסגת. ביברס ו מושקים ידועים בהיותם חצי-קווטיים,[1] אבל המכרסם המותאם ביותר לחיים ימיים הוא כנראה עכברוש מים ללא אוזניים מגינאה החדשה.[12] מכרסמים שגשגו גם בסביבות שנוצרו על ידי בני אדם כמו חַקלָאִי ו אזורים עירוניים.[13]

למרות שמינים מסוימים הם מזיקים שכיחים לבני אדם, מכרסמים ממלאים גם תפקידים אקולוגיים חשובים.[1] כמה מכרסמים נחשבים מיני אבן ו מהנדסי מערכות אקולוגיות בבתי הגידול שלהם. בתוך ה תוכניות גדולות בצפון אמריקה, פעילויות הנבירה של כלבי הערבה ממלאות תפקידים חשובים בקרקע אִוְרוּר וחלוקה מחדש של חומרים מזינים, העלאת התוכן האורגני בקרקע והגברת ספיגת המים. הם שומרים על בתי הגידול האלה,[14] וכמה אוכלי עשב גדולים כגון בִּיזוֹן ו שורש מעדיפים לרעות ליד מושבות כלבי ערבה בגלל איכות התזונה המוגברת של המספוא.[15]
השמדת כלבי הערבה יכולה לתרום גם לאזורית ולמקומית אובדן מגוון ביולוגי, מניעת זרע מוגברת והקמת ושיחים פולשניים והתפשטותם.[14] מכרסמים נוברים עלולים לאכול את גופי הפרי של פטריות ולהפיץ נבגים דרך הצואה שלהם, ובכך מאפשר לפטריות להתפזר ולהיווצר מערכות יחסים סימביוטיות עם שורשי הצמחים (שלרוב אינם יכולים לשגשג בלעדיהם). ככאלה, מכרסמים אלה עשויים למלא תפקיד בשמירה על יערות בריאים.[16]
באזורים ממוזגים רבים, הבונים משחקים חיוני הידרולוגי תַפְקִיד. בבניית הסכרים והלשכות שלהם, הבונים משנים את שבילי הנחלים והנהרות[17] ולאפשר יצירת בתי גידול נרחבים של ביצות. מחקר אחד מצא כי הנדסה על ידי בונים מובילה לגידול של 33 אחוזים במספר צמח עשבוני מינים ב אזורי מעבר.[18] מחקר אחר מצא כי בונים מגדילים את אוכלוסיות סלמון הבר.[19]
התנהגות ותולדות חיים
הַאֲכָלָה

רוב המכרסמים הם אוכלי עשב, ניזון אך ורק מחומר צמחי כגון זרעים, גבעולים, עלים, פרחים ושורשים. חלקם כן אוֹכֵל הַכָּל וכמה הם טורפים.[2] ה נשר שדה הוא מכרסם אוכלי עשב טיפוסי וניזון מעשבים, עשבי תיבול, פקעות שורש, אזוב וצמחייה אחרת, ומכרסם קליפות במהלך החורף. מדי פעם הוא אוכל חסרי חוליות כמו זחלי חרקים.[20] ה גופר בכיס המישורים אוכל חומר צמחי שנמצא מתחת לאדמה במהלך המנהרה, ואוסף גם עשבים, שורשים ופקעות בשקיות הלחיים שלו מטמונים אותם בתאי ספה תת קרקעיים.[21]
ה גופר כיס בטקסס נמנע מלהגיח אל פני השטח כדי להאכיל בכך שהוא תופס את שורשי הצמחים עם לסתותיו ומושך אותם מטה אל תוך מחילתו. זה גם מתרגל קופרופגיה.[22] ה חולדה שקופה אפריקאית מזונות על פני השטח, אוספים כל דבר שעשוי להיות אכיל לתוך שקיות הלחיים הגדולות שלו עד שפניו בולטות הצידה. לאחר מכן הוא חוזר למחילתו כדי למיין את החומר שאסף ואוכל את הפריטים המזינים.[23]
אגוטי מינים הם אחת מקבוצות החיות הבודדות שיכולות לפרוץ את הקפסולות הגדולות של אגוז ברזילאי פרי. יותר מדי זרעים נמצאים בפנים בכדי לצרוך אותם בארוחה אחת, ולכן האגוטי מנשא אותם ושומר אותם במטמון. זה עוזר בפיזור הזרעים שכן כל מה שהאגוטי לא מצליח לאחוז מרוחק מעץ האב כשהם נובטים. עצים אחרים נושאי אגוזים נוטים לשאת טיפת פירות בסתיו. אלה רבים מכדי לאכול אותם בארוחה אחת והסנאים אוספים ומאחסנים את העודפים בסדקים ובעצים חלולים. באזורים מדבריים, זרעים לרוב זמינים רק לתקופות קצרות. חולדת הקנגורו אוספת את כל מה שהיא מצליחה ומאחסנת אותם בתאי ספה במחילה שלה.[23]

אסטרטגיה להתמודדות עם שפע עונתי היא לאכול כמה שיותר ולאחסן את עודפי החומרים המזינים כשומן. מרמוטים עשה זאת ועשוי להיות כבד ב 50% בסתיו מאשר באביב. הם מסתמכים על מאגרי השומן שלהם במהלך החורף הארוך שלהם תַרְדֵמָה.[23] ביברס ניזונים מהעלים, הניצנים והקליפה הפנימית של עצים צומחים, כמו גם צמחי מים. הם מאחסנים אוכל לשימוש בחורף על ידי כריתת עצים קטנים וענפים עלים בסתיו וטבילתם בבריכתם, תוחבים את הקצוות בבוץ כדי לעגן אותם. כאן הם יכולים לגשת לאספקת המזון שלהם מתחת למים גם כאשר הבריכה שלהם קפואה.[24]
למרות שמכרסמים נחשבו באופן מסורתי ככלי עשב, מספר מינים כוללים בתזונה חרקים, דגים או בשר וצורות מיוחדות יותר מסתמכות על מאכלים כאלה. מחקר פונקציונלי-מורפולוגי של מערכת שן המכרסמים תומך ברעיון שמכרסמים פרימיטיביים היו אוכלי כל ולא אוכלי עשב. מחקרים בספרות מראים כי רבים מחברי ה- Sciuromorpha ו- Myomorpha, וכמה מבני ה- Hystricomorpha, כללו חומר בעלי חיים בתזונתם או שהיו מוכנים לאכול אוכל כזה כאשר הציעו אותו בשבי. בחינת תכולת הקיבה של צפון אמריקה עכבר לבן רגליים, שנחשב בדרך כלל כבעלי-עשב, הראה 34% חומר מן החי.[25]
טורפים מיוחדים יותר כוללים את חולדות ממולחות של הפיליפינים, הניזונים מחרקים וחסרי חוליות בעלי גוף רך, והאוסטרלים עכברוש מים, הזולל חרקים ימיים, דגים, סרטנים, מולים, חלזונות, צפרדעים, ביצי ציפורים ועופות מים.[25][26] ה עכבר חגב מאזורים יבשים בצפון אמריקה ניזון מחרקים, עקרבים ועכברים קטנים אחרים, ורק חלק קטן מהתזונה שלה הוא חומר צמחי. יש לו גוף שמנמן עם רגליים קצרות וזנב, אבל הוא זריז ויכול להכריע בקלות את הטרף כמו עצמו.[27]
התנהגות חברתית

מכרסמים מציגים מגוון רחב של סוגים של התנהגות חברתית החל מהיונק מערכת קסטות של ה חולדה עירומה,[28] ה"עיירה "הנרחבת של הקולוניאלי כלב ערבה,[29] דרך קבוצות משפחתיות לחיים העצמאיים והמתבודדים של בית מעונות למאכל. למעונות מבוגרים יש טווחי האכלה חופפים, אך הם חיים בקנים בודדים וניזונים בנפרד, ומתאחדים בקצרה בעונת הרבייה כדי להזדווג. ה גופר בכיס הוא גם חיה בודדה מחוץ לעונת הרבייה, כל אדם חופר מערכת מנהרות מורכבת ושומר על שטח.[9]
מכרסמים גדולים יותר נוטים להתגורר בתא משפחתי בו ההורים וצאצאיהם גרים יחד עד שהצעירים מתפזרים. ביברס לחיות ביחידות משפחתיות מורחבות, בדרך כלל עם זוג מבוגרים, ערכות השנה, צאצאי השנה הקודמת ולפעמים צעירים מבוגרים.[30] חולדות חומות בדרך כלל חיות במושבות קטנות עד שש נקבות חולקות מאורה וזכר אחד מגן על שֶׁטַח סביב המחילה. בצפיפות אוכלוסייה גבוהה, מערכת זו מתפרקת וגברים מראים מערכת היררכית של שְׁלִיטָה עם טווחים חופפים. צאצאים נשיים נשארים במושבה בזמן שצעירים גברים מתפזרים.[31] ה ערבה ערבה הוא מונוגמי ויוצר קשר זוגי לכל החיים. מחוץ לעונת הרבייה, שורשי ערבה חיים בסמיכות עם אחרים במושבות קטנות. זכר אינו תוקפני כלפי זכרים אחרים עד שהוא מזדווג, ולאחר מכן הוא מגן על טריטוריה, נקבה וקן נגד זכרים אחרים. הצמד מצטופף, מטפח זה בזה, וחולק אחריות קינון וגידול גור.[32]

בין המכרסמים החברתיים ביותר הם סנאי הקרקע, שנוצרים בדרך כלל מושבות מבוסס על קרבה נשית, כאשר גברים מתפזרים לאחר הגמילה והופכים נודדים כמבוגרים. שיתוף הפעולה בסנאים קרקעיים משתנה בין המינים וכולל בדרך כלל ביצוע שיחות אזעקה, הגנה על שטחים, שיתוף מזון, הגנה על אזורי קינון ומניעת רצח תינוקות.[33] ה כלב ערבה עם זנב שחור יוצר עיירות גדולות שעשויות לכסות דונמים רבים. המחילות אינן מקושרות זו בזו, אלא נחפרות ונכבשות על ידי קבוצות משפחתיות טריטוריאליות המכונות ערסים. קבוצה מורכבת לעיתים קרובות מזכר בוגר, שלוש או ארבע נקבות בוגרות, כמה בני נוער שאינם מגדלים, וצאצאי השנה הנוכחית. אנשים הנמצאים בקבוצות חביבות ידידותיים זה עם זה, אך עוינים כלפי זרים.[29]
אולי הדוגמאות הקיצוניות ביותר להתנהגות קולוניאלית אצל מכרסמים הן eusocial עכברוש שומה עירום עכברוש שומה דמראלנד. חולדת השומה העירומה חיה לגמרי מתחת לאדמה ויכולה ליצור מושבות של עד 80 פרטים. רק נקבה אחת ועד שלוש זכרים במושבה מתרבות, בעוד שאר החברים קטנים וסטריליים ומתפקדים כפועלים. ישנם אנשים בעלי גודל ביניים. הם עוזרים לגידול הצעירים ויכולים לתפוס את מקומו של רבייה אם מת.[28] חולדת השומה דמראלנד מאופיינת בכך שיש זכר ונקבה פעילים ורבייה במושבה שבה בעלי החיים הנותרים אינם סטריליים באמת, אלא הופכים לפוריים רק אם הם מקימים מושבה משלהם.[34]
תִקשׁוֹרֶת
חוש הריח

מכרסמים משתמשים בסימון ריחות בהקשרים חברתיים רבים, כולל תקשורת בין-פנים-מינים, סימון שבילים והקמת שטחים. השתן שלהם מספק מידע גנטי על פרטים, כולל המין, המין וזהות הפרט, ומידע מטבולי על דומיננטיות, מצב הרבייה ובריאות. תרכובות שמקורן ב מתחם תאימות היסטורי מרכזי (MHC) קשורים למספר חלבוני שתן. ריחו של טורף מדכא התנהגות של סימון ריח.[35]
מכרסמים מסוגלים לזהות קרובי משפחה לפי ריח וזה מאפשר להם להראות פּרוֹטֶקצִיָה (התנהגות מועדפת כלפי קרוביהם) וגם הימנעות מהכלאה. זֶה זיהוי קרובי משפחה הוא על ידי חוש הריח רמזים משתן, צואה והפרשות בלוטות. ההערכה העיקרית עשויה להיות כרוכה ב- MHC, כאשר מידת הקשר של שני אנשים מתואמת לגנים ה- MHC המשותפים להם. בתקשורת שאינם קרובי משפחה, שבהם נדרשים סמני ריח קבועים יותר, כמו בגבולות הטריטוריאליים, אז אינם נדיפים חלבונים עיקריים בשתן (MUP), המתפקדים כ- פרומון ניתן להשתמש גם במובילים. MUPs עשויים גם לסמן זהות פרטנית, עם כל זכר עכבר הבית (שריר מוס) מפריש שתן המכיל כתריסר MUPs מקודדים גנטית.[36]
עכברי בית מפקידים שתן המכיל פרומונים לצורך סימון טריטוריאלי, הכרה פרטנית וקבוצתית וארגון חברתי.[37] בונים טריטוריאליים סנאים אדומים לחקור ולהכיר את ניחוחות שכניהם ולהגיב בצורה פחות אגרסיבית לחדירות מצדם מאשר לאלה ש"שייטים "או זרים לא טריטוריאליים. זה מכונה "אפקט אויב יקר ".[38][39]
שְׁמִיעָתִי

למיני מכרסמים רבים, במיוחד כאלה שהם יומיים וחברתיים, יש מגוון רחב של שיחות אזעקה שנפלטות כאשר הם תופסים איומים. ישנם יתרונות ישירים ועקיפים של ביצוע פעולה זו. טורף פוטנציאלי עשוי להפסיק כאשר הוא יודע שהוא אותר, או שיחת אזעקה יכולה לאפשר מידע או אנשים קשורים לנקוט בפעולות התחמקות.[40] למספר מינים, למשל כלבי ערבה, יש מערכות שיחת אזעקה מורכבות נגד טורפים. למינים אלו עשויות להיות קריאות שונות לטורפים שונים (למשל טורפים מהאוויר או טורפים קרקעיים) וכל קריאה מכילה מידע על אופי האיום המדויק.[41] דחיפות האיום מועברת גם על ידי המאפיינים האקוסטיים של השיחה.[42]
למכרסמים חברתיים יש מגוון רחב יותר של קוליות מאשר למינים בודדים. 15 סוגים שונים של שיחות הוכרו אצל מבוגרים חולדות שומה קטאבה וארבעה אצל צעירים.[43] באופן דומה, ה degu משותף, מכרסם חברתי נוסף, מתחיל, מציג מגוון רחב של שיטות תקשורת ויש לו רפרטואר ווקאלי מורכב המורכב מחמש עשרה קטגוריות שונות של צליל.[44] לשיחות אולטרסאונד יש חלק בתקשורת החברתית בין מעונות ומשמשים אותם כאשר האינדיבידואלים לא נמצאים זה בזה.[45]
עכברי בית משתמשים בשיחות קוליות וקריאות אולטראסוניות במגוון הקשרים. לעתים קרובות ניתן לשמוע קולות נשמעים במהלך מפגשים אגוניסטיים או אגרסיביים, ואילו אולטרסאונד משמש בתקשורת מינית וגם אצל גורים כאשר הם נשרו מהקן.[37]

חולדות מעבדה (שהם חולדות חומות, Rattus norvegicus) פולטים קולות קצרים בתדירות גבוהה בקולי קולות במהלך חוויות מהנות כביכול כמו משחק מחוספס, כאשר צופים מינונים שגרתיים של מוֹרפִין, במהלך ההזדווגות, וכאשר מדגדגים. הושוו לווקאליזציה, שתוארה כ"ציוץ "מובהק צחוק, ומתפרש כציפייה למשהו מתגמל. במחקרים קליניים, הציוץ נקשר ברגשות רגשיים חיוביים, וקשר חברתי מתרחש עם הדגדוג, וכתוצאה מכך החולדות מתנות לחפש את הדגדוג. עם זאת, ככל שהחולדות מזדקנות, הנטייה לציוץ יורדת. כמו רוב קולות החולדה, הציוץ הוא בתדרים גבוהים מדי מכדי שבני אדם יוכלו לשמוע ללא ציוד מיוחד גלאי עטלפים שימשו למטרה זו.[46]
חָזוּתִי
מכרסמים, כמו כל יונקי השליה למעט פרימטים, יש רק שני סוגים של חרוטים קולטני אור ברשתית שלהם,[47] סוג "כחול-UV" אורך גל קצר וסוג "ירוק" של אורך גל אמצעי. לכן הם מסווגים כ דיכרומטים; עם זאת, הם רגישים ויזואלית לספקטרום האולטרה סגול (UV) ולכן הם יכולים לראות אור שבני אדם לא יכולים. הפונקציות של רגישות UV זו לא תמיד ברורות. ב degus, למשל, הבטן מחזירה יותר אור UV מאשר הגב. לכן, כאשר דגו קם על רגליו האחוריות, מה שהוא עושה כאשר נבהל, הוא חושף את בטנו לדיגומים אחרים וראייה אולטרה סגולה עשויה לשרת מטרה בהעברת האזעקה. כאשר הוא עומד על ארבע, גב החזרת ה- UV הנמוך שלו יכול לסייע בהפיכת הדגו פחות לטורפים.[48] אור אולטרה סגול נמצא בשפע ביום אך לא בלילה. יש עלייה גדולה ביחס בין אולטרה סגול לאור הנראה בשעות דמדומי הבוקר והערב. מכרסמים רבים פעילים בשעות הדמדומים (פעילות עורקית), ורגישות UV תהיה מועילה בזמנים אלה. רפלקטיביות אולטרה סגולה היא בעלת ערך מפוקפק עבור מכרסמים ליליים.[49]
השתן של מכרסמים רבים (למשל נברנים, דגים, עכברים, חולדות) מחזיר מאוד אור UV וזה יכול לשמש בתקשורת על ידי השארת סימני גלוי כמו גם ריח.[50] עם זאת, כמות ה- UV המשתקפת פוחתת עם הזמן, אשר בנסיבות מסוימות עלול להיות חסר; ה צואה מצוי יכול להבחין בין שבילי מכרסמים ישנים ורעננים וזוכה להצלחה גדולה יותר בציד במסלולים שסומנו לאחרונה.[51]
מִשׁוּשִׁי

רטט יכול לספק רמזים למושגים לגבי התנהגויות ספציפיות שמתבצעות, אזהרה והימנעות מטורפים, תחזוקת עדר או קבוצה וחיזור. ה חולדה חפרפרת עיוורת במזרח התיכון היה היונק הראשון שעבורו תקשורת סייסמית תועד. אלה פוסוריאלי מכרסמים דופקים את ראשם בקירות המנהרות שלהם. התנהגות זו התפרשה בתחילה כחלק מהתנהגות בניית המנהרה שלהם, אך בסופו של דבר הבינו כי הם מייצרים אותות סייסמיים מעוצבים זמנית לתקשורת למרחקים ארוכים עם חולדות שומה שכנות.[52]
שימוש בקליפות רגליים נרחב כאזהרת טורפים או פעולת הגנה. הוא משמש בעיקר מכרסמים מאובנים או חצי מאובנים.[53] חולדת הקנגורו עם זנב הדגל מייצרת כמה דפוסים מורכבים של חיפוי רגלי במספר הקשרים שונים, אחד מהם הוא כאשר הוא נתקל בנחש. חתימת הרגליים עשויה להזהיר את הצאצאים הסמוכים אך ככל הנראה משדרת שהחולדה דרוכה מדי להתקפה מוצלחת, ובכך למנוע את המרדף הדורסני של הנחש.[52][54] מספר מחקרים הצביעו על שימוש מכוון ב תנודות קרקע כאמצעי לתקשורת פנים ספציפית במהלך החיזור בין חולדת שומה קייפ.[55] דוחות רגליים היו מעורבים בתחרות גברים-גברים; הזכר הדומיננטי מציין את פוטנציאל החזקתו במשאבים על ידי תיפוף, ובכך ממזער את המגע הפיזי עם יריבים פוטנציאליים.[52]
אסטרטגיות הזיווג

מינים מסוימים של מכרסמים הם מונוגמיים, כאשר זכר ונקבה בוגרים יוצרים קיימא קשר זוגי. מונוגמיה יכולה לבוא בשתי צורות; מחויב ופקולטטיבי. במונוגמיה מחויבת, שני ההורים דואגים לצאצאים ומשחקים חלק חשוב בהישרדותם. זה קורה במינים כגון עכברים מקליפורניה, עכברי אולדפילד, חולדות ענק מלגאיות ובונים. במין זה, גברים בדרך כלל מזדווגים רק עם בני זוגם. בנוסף לטיפול מוגבר בצעירים, מונוגמיה מחויבת יכולה להועיל גם לגבר הבוגר מכיוון שהיא מקטינה את הסיכויים שלעולם לא למצוא בן זוג או להזדווג עם נקבה פורייה. במונוגמיה פקולטטיבית, הזכרים אינם מספקים טיפול הורי ישיר ונשארים עם נקבה אחת מכיוון שהם אינם יכולים לגשת לאחרים בגלל היותם מפוזרים במרחב. נראה כי שרקני הערבה הם דוגמא לצורה זו של מונוגמיה, כאשר גברים שומרים ומגנים על נקבות בסביבתן.[56]
ב פוליגיני מינים, גברים ינסו לנהל מונופול ולהזדווג עם מספר נקבות. כמו במונוגמיה, פוליגניה במכרסמים יכולה לבוא בשתי צורות; הגנה ולא הגנה. פוליגניה הגנתית כוללת גברים השולטים בשטחים המכילים משאבים המושכים נקבות. זה קורה בסנאים קרקעיים כמו מרמוטות צהובות כרס, סנאי קרקע בקליפורניה, סנאי קרקע קולומביאניים ו סנאי הקרקע של ריצ'רדסון. גברים עם טריטוריות ידועים כזכרים "תושבים" והנקבות שחיות בשטחים נקראות נקבות "תושבות". במקרה של מרמוטות, נראה כי גברים תושבים לא מאבדים אי פעם את שטחים ומנצחים תמיד מפגשים עם זכרים פולשים. ידוע כי מינים מסוימים מגנים באופן ישיר על נקבותיהן התושבות והקטטות שנוצרו עלולות להוביל לפציעה קשה. אצל מינים עם פוליגניה שאינה הגנה, גברים אינם טריטוריאליים ומשוטטים באופן נרחב בחיפוש אחר נקבות למונופול. זכרים אלה מקימים היררכיות דומיננטיות, כאשר הזכרים הגבוהים ביותר זוכים לגישה הנשית ביותר. זה קורה אצל מינים כמו סנאי הקרקע של בלדינג וכמה מיני סנאי עצים.[56]
הֶפקֵרוּת, בו גם גברים וגם נקבות מזדווגים עם מספר פרטנרים, מופיע גם אצל מכרסמים. במינים כמו עכבר לבן הרגליים, הנקבות יולדות המלטות בעלות אבהות מרובה. הפקרות מובילה לעלייה תחרות זרע ולגברים נוטים להיות אשכים גדולים יותר. בתוך ה סנאי קרקע קייפ, האשכים של הזכר יכולים להיות 20 אחוז מאורך ראש-גוף.[56] למספר מיני מכרסמים מערכות זיווג גמישות שיכולות להשתנות בין מונוגמיה, פוליגניה והפקרות.[56]
מכרסמים נשיים ממלאים תפקיד פעיל בבחירת בני זוגם. גורמים התורמים להעדפה נשית עשויים לכלול את הגודל, הדומיננטיות והיכולת המרחבית של הזכר.[57] בחולדות השומה העירומות החיוניות, נקבה יחידה מונופול זיווג לפחות משלושה זכרים.[28]
ברוב מיני המכרסמים, כגון חולדות חומות ועכברי בית, בִּיוּץ מתרחש במחזור רגיל ואילו אצל אחרים, כמו נשרים, זה כן המושרה על ידי הזדווגות. במהלך ההדבקה, גברים ממיני מכרסמים מפקידים א תקע זיווג בפתח איברי המין של הנקבה, הן למניעת דליפת זרע והן להגנה מפני זכרים אחרים המזרעים את הנקבה. נקבות יכולות להסיר את התקע ועשויות לעשות זאת באופן מיידי או לאחר מספר שעות.[57]
לידה והורות

גם מכרסמים עשויים להיוולד אלטריאלי (עיוור, חסר שיער ומפותח יחסית) או טרום חברתי (בעיקר פרווה, עיניים פקוחות ומפותחות למדי) תלוי במין. מצב אלטריאלי אופייני לסנאים ועכברים, ואילו המצב הטרום-חברתי מתרחש בדרך כלל אצל מינים כמו חזירי ים ודורבנים. נקבות עם צעירים ממגורים בדרך כלל בונות קנים משוכללים לפני שהן יולדות ומתחזקות אותן עד שצאצאיהן יהיו גָמוּל. הנקבה יולדת בישיבה או בשכיבה והצעירים מגיחים לכיוון אליו היא פונה. הילודים יוצאים לראשונה מהקן מספר ימים לאחר שפקחו את עיניהם ובהתחלה ממשיכים לחזור באופן קבוע. ככל שהם מתבגרים ומפותחים יותר, הם מבקרים בקן בתדירות נמוכה יותר ועוזבים לצמיתות כשהם נגמלים.[58]
במינים טרום-חברתיים האימהות משקיעות מעט בבניית קן וחלקן כלל אינן בונות קנים. הנקבה יולדת בעמידה והצעירים מגיחים מאחוריה. אמהות ממין זה שומרות על קשר עם צעיריהן הניידים ביותר עם שיחות קשר אימהיות. אף על פי שהם עצמאיים יחסית ונגמלים תוך מספר ימים, צעירים טרום-חברתיים עשויים להמשיך לאחות ולהיות מטופחים על ידי אימם. גודל המלטות של מכרסמים משתנה גם ונקבות עם המלטות קטנות מבלות יותר זמן בקן מאשר אלה עם המלטות גדולות יותר.[58]

מכרסמים של האם מספקים הן טיפול ישיר של ההורים, כגון סיעוד, טיפוח, שליפה והצטופפות, והן הורות עקיפה, כגון אחסון מזון, בניית קן והגנה על צאצאיהם.[58] אצל מינים חברתיים רבים, צעירים עשויים להיות מטופלים על ידי אנשים פרט להוריהם, מנהג המכונה כל ההורים אוֹ גידול שיתופי. ידוע שזה קורה בכלבי ערבה שחורים בזנב ובסנאי הקרקע של בלדינג, שם לאמהות יש קנים משותפים ומניקות צעירים שאינם קשורים יחד עם שלהם. יש שאלה כלשהי האם אמהות אלו יכולות להבחין בין צעירים שלהן. בתוך ה מארה פטגונית, צעירים ממוקמים גם במערכות קהילה, אך אמהות אינן מאפשרות לצעירים אחרים משלהם לאחות.[59]
רצח תינוקות קיים במספר מיני מכרסמים וניתן לתרגל אותם על ידי מין מבוגרים משני המינים. הוצעו כמה סיבות להתנהגות זו, כולל מתח תזונתי, תחרות במשאבים, הימנעות מניהול מוטעה של ההורים, ובמקרה של גברים, ניסיון להפוך את האם לקליטה מינית. הסיבה האחרונה נתמכת היטב ב פרימטים ו אריות אך פחות מכך במכרסמים.[60] נראה כי רצח תינוקות נפוץ בכלבי ערבה עם זנב שחור, כולל רצח תינוקות מזכרים פולשים ונקבות עולות, כמו גם מדי פעם קָנִיבָּלִיוּת של צאצאי הפרט עצמו.[61] כדי להגן מפני התינוקויות בקרב מבוגרים אחרים, מכרסמים עשויים להימנע מהתנפלות ישירה או נגד תוקפים פוטנציאליים, הזדווגות מרובה, טריטוריאליות או הפסקת הריון מוקדמת.[60] הרסנות יכול להתרחש גם בקרב מכרסמים; ב מרמוטות אלפיניות נקבות דומיננטיות נוטות לדכא את הרבייה של הכפופים על ידי היותן אנטגוניסטיות כלפיהן בזמן שהן בהריון. הלחץ שנוצר גורם להפלת העוברים.[62]
אינטליגנציה

מכרסמים התקדמו קוגניטיבי יכולות. הם יכולים ללמוד במהירות להימנע מפתיונות מורעלים, מה שמקשה עליהם להתמודד עם מזיקים.[1] חזירי ים יכולים ללמוד ולזכור מסלולים מורכבים לאוכל.[63] סנאים וחולדות קנגורו מסוגלים לאתר מטמונים של מזון על ידי זיכרון מרחבי, ולא רק לפי ריח.[64][65]
מכיוון שעכברי מעבדה (עכברי בית) וחולדות (חולדות חומות) נמצאים בשימוש נרחב כ מודלים מדעיים כדי לקדם את הבנתנו בביולוגיה, הרבה מאוד ידע על יכולותיהם הקוגניטיביות. חולדות חומות מציגות הטיה קוגניטיבית, כאשר עיבוד המידע מוטה על ידי האם הם במצב רגשי חיובי או שלילי.[66] לדוגמא, חולדות מעבדה שהוכשרו להגיב לטון ספציפי על ידי לחיצה על מנוף לקבלת תגמול, ולחיצה על מנוף אחר בתגובה לטון אחר בכדי למנוע התחשמלות, נוטות יותר להגיב לטון ביניים. על ידי בחירת מנוף התגמול אם זה עתה דגדג אותו (דבר שהם נהנים ממנו), תוך ציון "קשר בין המצב הרגשי החיובי שנמדד ישירות לבין קבלת החלטות בחוסר וודאות במודל חיה."[67]
לחולדות מעבדה (חומות) יכול להיות יכולת מטה-קוגניציה —to consider their own learning and then make decisions based on what they know, or do not know, as indicated by choices they make apparently trading off difficulty of tasks and expected rewards, making them the first animals other than primates known to have this capacity,[68][69] but these findings are disputed, since the rats may have been following simple התניה אופרנטית עקרונות,[70] או א behavioral economic דֶגֶם.[71] Brown rats use social learning in a wide range of situations, but perhaps especially so in acquiring food preferences.[72][73]
סיווג ואבולוציה
היסטוריה אבולוציונית

Dentition is the key feature by which מְאוּבָּן rodents are recognized and the earliest record of such mammals comes from the פליאוקן, shortly after the extinction of the non-avian דינוזאורים some 66 million years ago. These fossils are found in לוראסיה,[74] the supercontinent composed of modern-day North America, Europe, and Asia. The divergence of גלירס, a clade consisting of rodents and lagomorphs (rabbits, hares and pikas), from other placental mammals occurred within a few million years after the Cretaceous-Tertiary boundary; rodents and lagomorphs then הקרין במהלך קנוזואיק.[75] כמה שעון מולקולרי data suggest modern rodents (members of the order Rodentia) had appeared by the late קרטיקון, although other molecular divergence estimations are in agreement with the fossil record.[76][77]
Rodents are thought to have evolved in Asia, where local רב שכבתי faunas were severely affected by the אירוע הכחדה קרטיקון –פליאוגן and never fully recovered, unlike their North American and European relatives. In the resulting ecological vacuum, rodents and other Glires were able to evolve and diversify, taking the niches left by extinct multituberculates. The correlation between the spread of rodents and the demise of multituberculates is a controversial topic, not fully resolved. American and European multituberculate assemblages do decline in diversity in correlation with the introduction of rodents in these areas, but the remaining Asian multituberculates co-existed with rodents with no observable replacement taking place, and ultimately both clades co-existed for at least 15 million years.[78]
The history of the colonization of the world's continents by rodents is complex. The movements of the large superfamily מורואידאה (לְרַבּוֹת אוגרים, גרבילים, נכון עכברים ו חולדות ) may have involved up to seven colonizations of Africa, five of North America, four of Southeast Asia, two of South America and up to ten of Eurasia.[79]

במהלך אאוקן, rodents began to diversify. Beavers appeared in Eurasia in the late Eocene before spreading to North America in the late Miocene.[81] Late in the Eocene, hystricognaths invaded Africa, most probably having originated in Asia at least 39.5 million years ago.[82] From Africa, fossil evidence shows that some hystricognaths (caviomorphs ) colonized South America, which was an isolated continent at the time, evidently making use of ocean currents to cross the Atlantic on floating debris.[83] Caviomorphs had arrived in South America by 41 million years ago (implying a date at least as early as this for hystricognaths in Africa),[82] והיה reached the Greater Antilles על ידי המוקדם אוליגוקן, suggesting that they must have dispersed rapidly across South America.[84]
Nesomyid rodents are thought to have rafted from Africa to Madagascar 20–24 million years ago.[85] All 27 species of native Malagasy rodents appear to be descendants of a single colonization event.
By 20 million years ago, fossils recognizably belonging to the current families such as Muridae had emerged.[74] דרך מיוקן, when Africa had collided with Asia, African rodents such as the דָרְבָּן began to spread into אירואסיה.[86] Some fossil species were very large in comparison to modern rodents and included the giant beaver, Castoroides ohioensis, which grew to a length of 2.5 m (8 ft 2 in) and weight of 100 kg (220 lb).[87] The largest known rodent was Josephoartigasia monesi, א פקארנה with an estimated body length of 3 m (10 ft).[88]
The first rodents arrived in Australia via Indonesia around 5 million years ago. למרות ש בעלי חיים are the most prominent mammals in Australia, many מכרסמים, all belonging to the subfamily Murinae, are among the continent's mammal species.[89] There are about fifty species of 'old endemics', the first wave of rodents to colonize the country in the Miocene and early פליוקן, and eight true rat (ראטוס ) species of 'new endemics', arriving in a subsequent wave in the late Pliocene or early פליסטוקן. The earliest fossil rodents in Australia have a maximum age of 4.5 million years,[90] and molecular data is consistent with the colonization of New Guinea from the west during the late Miocene or early Pliocene followed by rapid diversification. A further wave of קרינה אדפטיבית occurred after one or more colonizations of Australia some 2 to 3 million years later.[91]
Rodents participated in the מחלף אמריקאי נהדר that resulted from the joining of the Americas by formation of the איסתמוס של פנמה, around 3 million years ago in the פיאצ'נזיאן גיל.[92] In this exchange, a small number of species such as the New World porcupines (Erethizontidae) headed north.[74] However, the main southward invasion of sigmodontines preceded formation of the land bridge by at least several million years, probably occurring via rafting.[93][94][95] Sigmodontines diversified explosively once in South America, although some degree of diversification may have already occurred in Central America before the colonization.[94][95] Their "head start" has relegated other North American rodent groups (sciurids, geomyids, heteromyids and nonsigmodontine cricetids) to a minor presence in the contemporary South American fauna.
Standard classification
השימוש ב- להזמין name "Rodentia" is attributed to the English traveler and naturalist תומאס אדוארד בודיץ ' (1821).[96] ה לטינית מודרנית מִלָה רודנטיה נגזר rodens, present participle of rodere – "to gnaw", "eat away".[97] ה ארנבות, ארנבות ו פיקאות (order Lagomorpha) have continuously growing incisors, as do rodents, and were at one time included in the order. However, they have an additional pair of incisors in the upper jaw and the two orders have quite separate evolutionary histories.[98] The phylogeny of the rodents places them in the לבושים Glires, Euarchontoglires ו בוריאוטריה. The cladogram below shows the inner and outer relations of Rodentia based on a 2012 attempt by Wu et al. to align the molecular clock with paleontological data:[99]
| בוריאוטריה |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||



The living rodent families based on the study done by Fabre et al. 2012.[100]



The order Rodentia may be divided into סדרי משנה, הפרות גבולות, משפחות על ו משפחות. יש הרבה מאוד מַקבִּילוּת ו הִתכַּנְסוּת among rodents caused by the fact that they have tended to evolve to fill largely similar niches. This parallel evolution includes not only the structure of the teeth, but also the infraorbital region of the skull (below the eye socket) and makes classification difficult as similar traits may not be due to common ancestry.[101][102] ברנדט (1855) was the first to propose dividing Rodentia into three suborders, Sciuromorpha, Hystricomorpha and Myomorpha, based on the development of certain muscles in the jaw and this system was widely accepted. Schlosser (1884) performed a comprehensive review of rodent fossils, mainly using the cheek teeth, and found that they fitted into the classical system, but Tullborg (1899) proposed just two sub-orders, Sciurognathi and Hystricognathi. These were based on the degree of inflection of the lower jaw and were to be further subdivided into Sciuromorpha, Myomorpha, Hystricomorpha and Bathyergomorpha. Matthew (1910) created a עץ פילוגנטי of New World rodents but did not include the more problematic Old World species. Further attempts at classification continued without agreement, with some authors adopting the classical three suborder system and others Tullborg's two suborders.[101]
These disagreements remain unresolved, nor have molecular studies fully resolved the situation though they have confirmed the מונופית of the group and that the clade has descended from a common Paleocene ancestor. Carleton and Musser (2005) in מינים של יונקים בעולם have provisionally adopted a five suborder system: Sciuromorpha, Castorimorpha, Myomorpha, Anomaluromorpha, and Hystricomorpha. These include 33 families, 481 genera and 2277 species.[103][104]

Order Rodentia (from Latin, rodere, to gnaw)





- סדר משנה אנומלורומורפה
- מִשׁפָּחָה אנומלורידים: scaly-tailed squirrels
- מִשׁפָּחָה פדטידה: springhares
- סדר משנה קסטורימורפה
- משפחה על קסטורואידאה
- מִשׁפָּחָה קסטורידים: beavers
- משפחה על Geomyoidea
- מִשׁפָּחָה Geomyidae: pocket gophers (true gophers)
- מִשׁפָּחָה Heteromyidae: kangaroo rats, kangaroo mice
- משפחה על קסטורואידאה
- סדר משנה היסטריקומורפה
- אינפראורדר Ctenodactylomorphi
- מִשׁפָּחָה Ctenodactylidae: gundis
- אינפראורדר Hystricognathi
- מִשׁפָּחָה Bathyergidae: African mole rats
- מִשׁפָּחָה Hystricidae: Old World porcupines
- מִשׁפָּחָה Petromuridae: dassie rat
- מִשׁפָּחָה Thryonomyidae: cane rats
- מזגן Caviomorpha
- משפחה †Heptaxodontidae: giant hutias
- מִשׁפָּחָה Abrocomidae: chinchilla rats
- מִשׁפָּחָה Capromyidae: hutias
- מִשׁפָּחָה Caviidae: cavies, including שרקנים וה קפיברה
- מִשׁפָּחָה צ'ינצ'ילידים: צ'ינצ'ילות, ויסקצ'ות
- מִשׁפָּחָה Ctenomyidae: טוקו-טוקו
- מִשׁפָּחָה Dasyproctidae: agoutis
- מִשׁפָּחָה Cuniculidae: pacas
- מִשׁפָּחָה דומיניות: pacaranas
- מִשׁפָּחָה Echimyidae: spiny rats
- מִשׁפָּחָה Erethizontidae: New World porcupines
- מִשׁפָּחָה מיוקסטורידים: coypu (nutria)
- מִשׁפָּחָה אוקטודונטיות: octodonts
- אינפראורדר Incertae sedis
- מִשׁפָּחָה Diatomyidae: Laotian rock rat
- אינפראורדר Ctenodactylomorphi
- סדר משנה מיומורפה
- משפחה על Dipodoidea
- מִשׁפָּחָה Dipodidae: jerboas and jumping mice
- משפחה על מורואידאה
- מִשׁפָּחָה Calomyscidae: mouse-like hamsters
- מִשׁפָּחָה חרסנים: אוגרים, חולדות ועכברים חדשים של העולם, muskrats, voles, lemmings
- מִשׁפָּחָה Muridae: true mice and rats, gerbils, spiny mice, crested rat
- מִשׁפָּחָה Nesomyidae: climbing mice, rock mice, white-tailed rat, Malagasy rats and mice
- מִשׁפָּחָה Platacanthomyidae: spiny dormice
- מִשׁפָּחָה Spalacidae: mole rats, bamboo rats, zokors
- משפחה על Dipodoidea
- סדר משנה Sciuromorpha
- מִשׁפָּחָה Aplodontiidae: mountain beaver
- מִשׁפָּחָה Gliridae (also Myoxidae, Muscardinidae): dormice
- מִשׁפָּחָה Sciuridae: squirrels, including שבבי שבבים, prairie dogs, marmots
אינטראקציה עם בני אדם
שימור

While rodents are not the most seriously threatened order of mammals, there are 168 species in 126 genera that are said to warrant conservation attention[105] in the face of limited appreciation by the public. Since 76 percent of rodent genera contain only one species, much phylogenetic diversity could be lost with a comparatively small number of extinctions. In the absence of more detailed knowledge of species at risk and accurate taxonomy, conservation must be based mainly on higher taxa (such as families rather than species) and geographical hot spots.[105] כמה מינים של rice rat have become extinct since the 19th century, probably through habitat loss and the introduction of alien species.[106] In Colombia, the brown hairy dwarf porcupine was recorded from only two mountain localities in the 1920s, while the red crested soft-furred spiny rat is known only from its סוג יישוב on the Caribbean coast, so these species are considered vulnerable.[107] The IUCN Species Survival Commission writes "We can safely conclude that many South American rodents are seriously threatened, mainly by environmental disturbance and intensive hunting".[108]
The "three now cosmopolitan commensal rodent pest species"[109] (the brown rat, the חולדה שחורה and the house mouse) have been dispersed in association with humans, partly on sailing ships in the עידן החקר, and with a fourth species in the Pacific, the Polynesian rat (Rattus exulans), have severely damaged island biotas מסביב לעולם. For example, when the black rat reached האי לורד האו in 1918, over 40 percent of the terrestrial bird species of the island, including the Lord Howe fantail,[110] became extinct within ten years. Similar destruction has been seen on האי מידוויי (1943) ו האי דרום קייפ הגדול (1962). Conservation projects can with careful planning completely eradicate these pest rodents from islands using an נוגד קרישה rodenticide such as brodifacoum.[109] This approach has been successful on the island of לונדי in the United Kingdom, where the eradication of an estimated 40,000 brown rats is giving populations of מי גזירה מאנקס ו פופין אטלנטי a chance to recover from near-extinction.[111][112]
ניצול

Humanity has long used animal skins for clothing, as the leather is durable and the פרווה provides extra insulation.[2] The native people of North America made much use of beaver pelts, tanning and sewing them together to make robes. Europeans appreciated the quality of these and the סחר בפרווה בצפון אמריקה developed and became of prime importance to early settlers. In Europe, the soft underfur known as "beaver wool" was found to be ideal for felting and was made into beaver hats and trimming for clothing.[113][114] מאוחר יותר, ה קויפו took over as a cheaper source of fur for felting and was farmed extensively in America and Europe; however, fashions changed, new materials became available and this area of the animal fur industry declined.[115] ה צ'ינצ'ילה has a soft and silky coat and the demand for its fur was so high that it was nearly wiped out in the wild before farming took over as the main source of pelts.[115] The quills and guardhairs of porcupines are used for traditional decorative clothing. For example, their guardhairs are used in the creation of the יליד אמריקאי "porky roach" headdress. The main quills may be dyed, and then applied in combination with thread to embellish leather accessories such as knife sheaths and leather bags. לקוטה women would harvest the quills for quillwork by throwing a blanket over a porcupine and retrieving the quills it left stuck in the blanket.[116]
צְרִיכָה
At least 89 species of rodent, mostly Hystricomorpha such as guinea pigs, agoutis and capybaras, are eaten by humans; in 1985, there were at least 42 different societies in which people eat rats.[117] Guinea pigs were first raised for food around 2500 B.C. and by 1500 B.C. had become the main source of meat for the אימפריית האינקה. Dormice were raised by the Romans in special pots called "gliraria", or in large outdoor enclosures, where they were fattened on walnuts, chestnuts, and acorns. The dormice were also caught from the wild in autumn when they were fattest, and either roasted and dipped into honey or baked while stuffed with a mixture of pork, pine nuts, and other flavorings. Researchers found that in Amazonia, where large mammals were scarce, pacas and common agoutis accounted for around 40 percent of the annual game taken by the indigenous people, but in forested areas where larger mammals were abundant, these rodents constituted only about 3 percent of the take.[117]
Guinea pigs are used in the cuisine of קוזקו, Peru, in dishes such as cuy al horno, baked guinea pig.[2][118] The traditional Andean stove, known as a qoncha או א fogón, is made from mud and clay reinforced with straw and hair from animals such as guinea pigs.[119] In Peru, there are at any time 20 million domestic guinea pigs, which annually produce 64 million edible carcasses. This animal is an excellent food source since the flesh is 19% protein.[117] In the United States, mostly squirrels, but also muskrats, porcupines, and גידולי קרקע are eaten by humans. ה אנשי נאוואחו ate prairie dog baked in mud, while the פאיוט ate gophers, squirrels, and rats.[117]
ניסויים בבעלי חיים

Rodents are used widely as אורגניזמים מודליים in animal testing.[2][120] Albino mutant חולדות were first used for research in 1828 and later became the first animal מְבוּיָת for purely scientific purposes.[121] Nowadays, the house mouse is the most commonly used laboratory rodent, and in 1979 it was estimated that fifty million were used annually worldwide. They are favored because of their small size, fertility, short gestation period and ease of handling and because they are susceptible to many of the conditions and infections that afflict humans. They are used in research into גנטיקה, ביולוגיה התפתחותית, ביולוגיה של התא, אונקולוגיה ו תוֹרַת הַחִסוּן.[122] Guinea pigs were popular laboratory animals until the late 20th century; about 2.5 million guinea pigs were used annually in the United States for research in the 1960s,[123] but that total decreased to about 375,000 by the mid-1990s.[124] In 2007, they constituted about 2% of all laboratory animals.[123] Guinea pigs played a major role in the establishment of תורת הנבט in the late 19th century, through the experiments of לואיס פסטר, אמיל רוקס, ו רוברט קוך.[125] הם היו launched into orbital space flight several times—first by the ברית המועצות על Sputnik 9 biosatellite of March 9, 1961, with a successful recovery.[126] The naked mole rat is the only known mammal that is poikilothermic; it is used in studies on ויסות תרמי. It is also unusual in not producing the neurotransmitter חומר P, a fact which researchers find useful in studies on כְּאֵב.[127]
Rodents have sensitive olfactory abilities, which have been used by humans to detect odors or chemicals of interest.[128] ה Gambian pouched rat is able to detect שַׁחֶפֶת bacilli with a sensitivity of up to 86.6%, and specificity (detecting the absence of the bacilli) of over 93%; the same species has been trained to detect מוקשים יבשתיים.[129][130] Rats have been studied for possible use in hazardous situations such as in disaster zones. They can be trained to respond to commands, which may be given remotely, and even persuaded to venture into brightly lit areas, which rats usually avoid.[131][132][133]
כחיות מחמד
Rodents including guinea pigs,[134] mice, rats, hamsters, gerbils, chinchillas, degus and chipmunks make convenient pets able to live in small spaces, each species with its own qualities.[135] Most are normally kept in cages of suitable sizes and have varied requirements for space and social interaction. If handled from a young age, they are usually docile and do not bite. Guinea pigs have a long lifespan and need a large cage.[63] Rats also need plenty of space and can become very tame, can learn tricks and seem to enjoy human companionship. Mice are short-lived but take up very little space. Hamsters are solitary but tend to be nocturnal. They have interesting behaviors, but unless handled regularly they may be defensive. Gerbils are not usually aggressive, rarely bite and are sociable animals that enjoy the company of humans and their own kind.[136]
As pests and disease vectors

Some rodent species are serious agricultural מזיקים, eating large quantities of food stored by humans.[137] For example, in 2003, the amount of אורז lost to mice and rats in Asia was estimated to be enough to feed 200 million people. Most of the damage worldwide is caused by a relatively small number of species, chiefly rats and mice.[138] In Indonesia and Tanzania, rodents reduce crop yields by around fifteen percent, while in some instances in South America losses have reached ninety percent. Across Africa, rodents including מאסטומי ו Arvicanthis damage cereals, groundnuts, vegetables and cacao. In Asia, rats, mice and species such as Microtus brandti, Meriones unguiculatus ו Eospalax baileyi damage crops of rice, sorghum, tubers, vegetables and nuts. In Europe, as well as rats and mice, species of אפודמוס, מיקרוטוס and in occasional outbreaks ארוויקולה טרסטריס cause damage to orchards, vegetables and pasture as well as cereals. In South America, a wider range of rodent species, such as הולוכילוס, אקודון, Calomys, אוליגוריזומיס, פילוטיס, Sigmodon ו Zygodontomys, damage many crops including sugar cane, fruits, vegetables, and tubers.[138]
Rodents are also significant vectors of disease.[139] The black rat, with the fleas that it carries, plays a primary role in spreading the bacterium Yersinia pestis אחראי ל מגפת דבר,[140] and carries the organisms responsible for טִיפוּס, Weil's disease, טוקסופלזמוזיס ו trichinosis.[139] A number of rodents carry hantaviruses, כולל ה פומואלה, דוברבה ו Saaremaa viruses, which can infect humans.[141] Rodents also help to transmit diseases including בייביוזיס, cutaneous לישמניאזיס, human granulocytic anaplasmosis, מחלת ליים, קדחת המורגי של אומסק, וירוס Powassan, אבעבועות, חום חוזר, הר הרוקי הבחין בקדחת, ו וירוס הנילוס המערבי.[142]

Because rodents are a nuisance and endanger בריאות ציבור, human societies often attempt to control them. Traditionally, this involved poisoning and trapping, methods that were not always safe or effective. יותר לאחרונה, הדברה משולבת attempts to improve control with a combination of surveys to determine the size and distribution of the pest population, the establishment of tolerance limits (levels of pest activity at which to intervene), interventions, and evaluation of effectiveness based on repeated surveys. Interventions may include education, making and applying laws and regulations, modifying the habitat, changing farming practices, and בקרה ביולוגית באמצעות פתוגנים אוֹ חיות טרף, בנוסף ל הַרעָלָה and trapping.[143] The use of pathogens such as סלמונלה has the drawback that they can infect man and domestic animals, and rodents often become resistant. The use of predators including חמוסים, נמיות ו לפקח לטאות has been found unsatisfactory. Domestic and feral חתולים are able to control rodents effectively, provided the rodent population is not too large.[144] In the UK, two species in particular, the house mouse and the brown rat, are actively controlled to limit damage in growing crops, loss and contamination of stored crops and structural damage to facilities, as well as to comply with the law.[145]
ראה גם
- Fe, Fi, Fo, Fum, and Phooey, mice who orbited the Moon on אפולו 17
- Mouse models of breast cancer metastasis
הפניות
- ^ א ב ג ד ה f ז ח Single, G.; Dickman, C. R.; MacDonald, D. W. (2001). "Rodents". In MacDonald, D. W. (ed.). האנציקלופדיה של היונקים (מהדורה שנייה). הוצאת אוניברסיטת אוקספורד. pp. 578–587. ISBN 978-0-7607-1969-5.
- ^ א ב ג ד ה f Waggoner, Ben (15 August 2000). "Introduction to the Rodentia". המוזיאון לפליאונטולוגיה של אוניברסיטת קליפורניה. הוחזר 4 ביולי 2014.
- ^ א ב ג ד ה f Nowak, R. M. (1999). יונקי העולם של ווקר. הוצאת אוניברסיטת ג'ונס הופקינס. עמ ' 1244. ISBN 978-0-8018-5789-8.
- ^ Blashfield, Jean F. "Rodents". Science Encyclopedia. הוחזר 14 ביולי 2014.
- ^ Niemiec, Brook A. (15 October 2011). Small Animal Dental, Oral and Maxillofacial Disease: A Colour Handbook. עיתונות CRC. עמ ' 13. ISBN 978-1-84076-630-1.
- ^ Cox, Philip G.; Jeffery, Nathan (2011). "Reviewing the Morphology of the Jaw-Closing Musculature in Squirrels, Rats, and Guinea Pigs with Contrast-Enhanced MicroCT" (PDF). הרשומה האנטומית. 294 (6): 915–928. דוי:10.1002/ar.21381. PMID 21538924. S2CID 17249666.
- ^ Duckett, W. (1853). "cheek pouch". English conversation and reading. Ed Michel Levi. עמ ' 3.
- ^ Mustapha, O. (2015). "Morphology of the Oral Cavity of the African Giant Rat". Bulgarian Journal of Veterinary Medicine. 18 (1): 19–30. דוי:10.15547/bjvm.793.
- ^ א ב ג Stefoff, Rebecca (2008). The Rodent Order. מרשל קוונדיש. pp. 62–63, 71–73. ISBN 978-0-7614-3073-5.
- ^ Thorington, R. W Jr.; Darrow, K.; Anderson, C. G. (1998). "Wing tip anatomy and aerodynamics in flying squirrels" (PDF). כתב העת לממלוגיה. 79 (1): 245–250. דוי:10.2307/1382860. JSTOR 1382860.
- ^ Schulte-Hostedde, A. I. (2008). "Chapter 10: Sexual Size Dimorphism in Rodents". In Wolff, Jerry O.; Sherman, Paul W. (eds.). Rodent Societies: An Ecological and Evolutionary Perspective. הוצאת אוניברסיטת שיקגו. עמ '117–119. ISBN 978-0-226-90538-9.
- ^ Helgen, Kristofer M. (2005). "The amphibious murines of New Guinea (Rodentia, Muridae): the generic status of בייאנקמיס and description of a new species of הידרומי". זואטקסה. 913: 1–20. דוי:10.11646/zootaxa.913.1.1. ISSN 1175-5326.
- ^ Parshad, V.R. (1999). "Rodent control in India" (PDF). ביקורות משולבות על הדברה. 4 (2): 97–126. דוי:10.1023/A:1009622109901. S2CID 36804001.
- ^ א ב ג'נקה, אקסל; Martínez-Estévez, Lourdes; Balvanera, Patricia; Pacheco, Jesús; Ceballos, Gerardo (2013). "Prairie dog decline reduces the supply of ecosystem services and leads to desertification of semiarid grasslands". PLOS ONE. 8 (10): e75229. ביבקוד:2013PLoSO...875229M. דוי:10.1371/journal.pone.0075229. ISSN 1932-6203. PMC 3793983. PMID 24130691.
- ^ Krueger, Kirsten (1986). "Feeding relationships among bison, pronghorn, and prairie dogs: an experimental analysis". אֵקוֹלוֹגִיָה. 67 (3): 760–770. דוי:10.2307/1937699. ISSN 0012-9658. JSTOR 1937699.
- ^ Pérez, Francisco; קסטילו-גווארה, סיטללי; Galindo-Flores, Gema; Cuautle, Mariana; Estrada-Torres, Arturo (2012). "Effect of gut passage by two highland rodents on spore activity and mycorrhiza formation of two species of ectomycorrhizal fungi (Laccaria trichodermophora ו Suillus tomentosus)". בּוֹטָנִיקָה. 90 (11): 1084–1092. דוי:10.1139/b2012-086. ISSN 1916-2790.
- ^ Burchsted, D.; דניאלס, מ '; Thorson, R.; Vokoun, J. (2010). "The river discontinuum: applying beaver modifications to baseline conditions for restoration of forested headwaters". BioScience. 60 (11): 908–922. דוי:10.1525/bio.2010.60.11.7. S2CID 10070184.
- ^ רייט, ג'יי פ '; Jones, C. G.; Flecker, A. S. (2002). "An ecosystem engineer, the beaver, increases species richness at the landscape scale" (PDF). אקולוגיה. 132 (1): 96–101. ביבקוד:2002Oecol.132...96W. דוי:10.1007/s00442-002-0929-1. PMID 28547281. S2CID 5940275.
- ^ Kemp, P. S.; Worthington, T. A.; Langford, T. E. l.; Tree, A. R. J.; Gaywood, M. J. (2012). "Qualitative and quantitative effects of reintroduced beavers on stream fish". דגים ודייג. 13 (2): 158–181. דוי:10.1111/j.1467-2979.2011.00421.x.
- ^ Hansson, Lennart (1971). "Habitat, food and population dynamics of the field vole מיקרוטוס אגרסטיס (L.) in south Sweden". Viltrevy. 8: 268–278. ISSN 0505-611X. הועבר לארכיון מ המקורי ב- 27 בספטמבר 2013.
- ^ Connior, M. B. (2011). "בוריס גאומי (Rodentia: Geomyidae)". מינים של יונקים. 43 (1): 104–117. דוי:10.1644/879.1.
- ^ "Texan pocket gopher". The Mammals of Texas: Rodents. NSRL: Museum of Texas Tech University. הוחזר 4 ביולי 2014.
- ^ א ב ג אטנבורו, דייוויד (2002). חיי היונקים. ספרי BBC. pp. 61–86. ISBN 978-0-563-53423-5.
- ^ Müller-Schwarze, Dietland; Sun, Lixing (2003). הבונה: היסטוריה טבעית של מהנדס שדות מים. הוצאת אוניברסיטת קורנל. pp. 67–75. ISBN 978-0-8014-4098-4.
- ^ א ב Landry, Stuart O. Jr. (1970). "The Rodentia as omnivores". הסקירה הרבעונית של הביולוגיה. 45 (4): 351–372. דוי:10.1086/406647. JSTOR 2821009. PMID 5500524. S2CID 30382320.
- ^ "Hydrysys chrysogaster: Water rat". Water for a healthy country. CSIRO. 30 ביוני 2004. הוחזר 4 ביולי 2014.
- ^ "Northern grasshopper mouse". The Mammals of Texas: Rodents. NSRL: Museum of Texas Tech University. הוחזר 4 ביולי 2014.
- ^ א ב ג Jarvis, Jennifer (1981). "Eusociality in a mammal: Cooperative breeding in naked mole-rat colonies". מַדָע. 212 (4494): 571–573. ביבקוד:1981Sci...212..571J. דוי:10.1126/science.7209555. JSTOR 1686202. PMID 7209555.
- ^ א ב Hoogland, John L. (1995). The Black-Tailed Prairie Dog: Social Life of a Burrowing Mammal. הוצאת אוניברסיטת שיקגו. עמ ' 1. ISBN 978-0-226-35118-6.
- ^ Baker, Bruce W.; Hill, Edward P. (2003). "Chapter 15: Beaver". In Feldhamer, George A.; Thompson, Bruce C.; Chapman, Joseph A. (eds.). יונקים פרועים של צפון אמריקה: ביולוגיה, ניהול ושימור. לחץ על JHU. pp. 288–310. ISBN 978-0-8018-7416-1.
- ^ Hanson, Anne (25 October 2006). "Wild Norway rat behavior". Rat behavior and biology. הוחזר 1 ביולי 2014.
- ^ Winslow, James T.; Hastings, Nick; Carter, C. Sue; Harbaugh, Carroll R.; Insel, Thomas R. (1993). "A role for central vasopressin in pair bonding in monogamous prairie voles" (PDF). Letters to Nature. 365 (6446): 545–548. ביבקוד:1993Natur.365..545W. דוי:10.1038/365545a0. PMID 8413608. S2CID 4333114. הועבר לארכיון מ המקורי (PDF) ב- 14 ביולי 2014.
- ^ Yensen, Eric; Sherman, Paul W. (2003). "Chapter 10: Ground Squirrels". In Feldhamer, George A.; Thompson, Bruce C.; Chapman, Joseph A. (eds.). יונקים פרועים של צפון אמריקה: ביולוגיה, ניהול ושימור. לחץ על JHU. pp. 211–225. ISBN 978-0-8018-7416-1.
- ^ בנט, נ 'ג'; Jarvis, J. U. M. (2004). "Cryptomys damarensis". מינים של יונקים. 756: Number 756: pp. 1–5. דוי:10.1644/756.
- ^ Arakawa, Hiroyuki; Blanchard, D. Caroline; Arakawa, Keiko; Dunlap, Christopher; Blanchard, Robert J. (2008). "Scent marking behavior as an odorant communication in mice". מדעי המוח וביקורות ביולוגיות התנהגותיות. 32 (7): 1236–1248. דוי:10.1016/j.neubiorev.2008.05.012. PMC 2577770. PMID 18565582.
- ^ Holmes, Warren G.; Mateo, Jill M. (2008). "Chapter 19: Kin Recognition in Rodents: Issues and Evidence". In Wolff, Jerry O.; Sherman, Paul W. (eds.). Rodent Societies: An Ecological and Evolutionary Perspective. הוצאת אוניברסיטת שיקגו. pp. 216–230. ISBN 978-0-226-90538-9.
- ^ א ב Sherwin, C.M. (2002). "Comfortable quarters for mice in research institutions". In Viktor and Annie Reinhardt (ed.). Comfortable Quarters For Laboratory Animals (מהדורה 9). Animal Welfare Institute. הועבר לארכיון מ המקורי ב- 6 באוקטובר 2014.
- ^ Bjorkoyli, Tore; Rosell, Frank (2002). "A test of the dear enemy phenomenon in the Eurasian beaver". התנהגות בעלי חיים. 63 (6): 1073–1078. דוי:10.1006/anbe.2002.3010. hdl:11250/2437993. S2CID 53160345.
- ^ Vaché, M.; Ferron, J.; Gouat, P. (2001). "The ability of red squirrels (Tamiasciurus hudsonicus) to discriminate conspecific olfactory signatures". כתב העת הקנדי לזואולוגיה. 79 (7): 1296–1300. דוי:10.1139/z01-085. S2CID 86280677.
- ^ Shelley, Erin L.; Blumstein, Daniel T. (2005). "The evolution of vocal alarm communication in rodents". אקולוגיה התנהגותית. 16 (1): 169–177. CiteSeerX 10.1.1.541.4408. דוי:10.1093/beheco/arh148.
- ^ Slobodchikoff, C. N.; Paseka, Andrea; Verdolin, Jennifer L (2009). "Prairie dog alarm calls encode labels about predator colors" (PDF). הכרת בעלי חיים. 12 (3): 435–439. דוי:10.1007/s10071-008-0203-y. PMID 19116730. S2CID 13178244.
- ^ Zimmermann, Elke; Leliveld, Lisette; Schehka, Lisette (2013). "8: Toward the evolutionary roots of affective prosody in human acoustic communication: A comparative approach to mammalian voices". In Altenmüller, Eckart; Schmidt, Sabine; Zimmermann, Elke (eds.). The Evolution of Emotional Communication: From Sounds in Nonhuman Mammals to Speech and Music in Man. הוצאת אוניברסיטת אוקספורד. עמ '123–124. ISBN 978-0-19-164489-4.
- ^ Vanden Hole, Charlotte; Van Daele, Paul A. A. G.; Desmet, Niels; Devos, Paul & Adriaens, Dominique (2014). "Does sociality imply a complex vocal communication system? A case study for Fukomys micklemi (Bathyergidae, Rodentia)". ביו-אקוסטיקה. 23 (2): 143–160. דוי:10.1080/09524622.2013.841085. S2CID 84503870.
- ^ Long, C. V. (2007). "Vocalisations of the degu (אוקטודון דגוס), a social caviomorph rodent". ביו-אקוסטיקה. 16 (3): 223–244. דוי:10.1080/09524622.2007.9753579. ISSN 0952-4622. S2CID 84569309.
- ^ Ancillotto, Leonardo; Sozio, Giulia; Mortelliti, Alessio; Russo, Danilo (2014). "Ultrasonic communication in Gliridae (Rodentia): the hazel dormouse (Muscardinus avellanarius) as a case study". ביו-אקוסטיקה. 23 (2): 129–141. דוי:10.1080/09524622.2013.838146. S2CID 84012458.
- ^ Panksepp, Jaak; Burgdorf, Jeff (2003). ""Laughing" rats and the evolutionary antecedents of human joy?". פיזיולוגיה והתנהגות. 79 (3): 533–547. CiteSeerX 10.1.1.326.9267. דוי:10.1016/S0031-9384(03)00159-8. PMID 12954448. S2CID 14063615.
- ^ Haverkamp, Silke; Waessle, Heinz; Duebel, Jens; Kuner, Thomas; Augustine, George J.; Feng, Guoping; Euler, Thomas (2005). "The primordial, blue-cone color system of the mouse retina". כתב העת למדעי המוח. 25 (22): 5438–5445. דוי:10.1523/JNEUROSCI.1117-05.2005. PMC 6725002. PMID 15930394.
- ^ Hanson, Anne. "What do rats see?". Rat behavior and biology. הוחזר 1 ביולי 2014.
- ^ Pickrell, John (8 July 2003). "Urine vision? How rodents communicate with UV light". חדשות נשיונל ג'יאוגרפיק. הוחזר 8 ביולי 2014.
- ^ Desjardins, C.; Maruniak, J. A.; Bronson, F. H. (1973). "Social rank in house mice: Differentiation revealed by ultraviolet visualization of urinary marking patterns". מַדָע. 182 (4115): 939–941. ביבקוד:1973Sci...182..939D. דוי:10.1126/science.182.4115.939. PMID 4745598. S2CID 44346136.
- ^ Viitala, J.; Korpimäki, E.; Palokangas, P.; Koivula, M. (1995). "Attraction of kestrels to vole scent marks visible in ultraviolet light". טֶבַע. 373 (6513): 425–427. ביבקוד:1995Natur.373..425V. דוי:10.1038/373425a0. S2CID 4356193.
- ^ א ב ג "Vibrational communication in mammals". Map of Life: Convergent evolution online. אוניברסיטת קמברידג. 4 באוגוסט 2010. הוחזר 5 ביולי 2014.
- ^ Randall, J. A. (2001). "Evolution and function of drumming as communication in mammals". זואולוג אמריקאי. 41 (5): 1143–1156. דוי:10.1093/icb/41.5.1143.
- ^ Randall, Jan A.; Matocq, Marjorie D. (1997). "Why do kangaroo rats (Dipodomys spectabilis) footdrum at snakes?". אקולוגיה התנהגותית. 8 (4): 404–413. דוי:10.1093/beheco/8.4.404.
- ^ Narins, P. M.; Reichman, O. J.; Jarvis, J. U. M.; Lewis, E. R. (1992). "Seismic signal transmission between burrows of the Cape mole-rat Georychus capensis". Journal of Comparative Physiology [A]. 170 (1): 13–22. דוי:10.1007/BF00190397. PMID 1573567. S2CID 22600955.
- ^ א ב ג ד Waterman, Jane (2008). "Chapter 3: Male Mating Strategies in Rodents". In Wolff, Jerry O.; Sherman, Paul W. (eds.). Rodent Societies: An Ecological and Evolutionary Perspective. הוצאת אוניברסיטת שיקגו. pp. 28–39. ISBN 978-0-226-90538-9.
- ^ א ב Soloman, Nancy G.; Keane, Brain (2008). "Chapter 4: Reproductive Strategies in Female Rodents". In Wolff, Jerry O.; Sherman, Paul W. (eds.). Rodent Societies: An Ecological and Evolutionary Perspective. הוצאת אוניברסיטת שיקגו. pp. 42–52. ISBN 978-0-226-90538-9.
- ^ א ב ג McGuire, Betty; Bernis, William E. (2008). "Chapter 20: Parental Care". In Wolff, Jerry O.; Sherman, Paul W. (eds.). Rodent Societies: An Ecological and Evolutionary Perspective. הוצאת אוניברסיטת שיקגו. pp. 231–235. ISBN 978-0-226-90538-9.
- ^ Holmes, Warren G.; Mateo, Jill M. (2008). "Chapter 19: Kin Recognition in Rodents: Issues and Evidence". In Wolff, Jerry O.; Sherman, Paul W. (eds.). Rodent Societies: An Ecological and Evolutionary Perspective. הוצאת אוניברסיטת שיקגו. pp. 226–227. ISBN 978-0-226-90538-9.
- ^ א ב Ebensperger, Luis A.; Blumsperger, Daniel T. (2008). "Chapter 23: Nonparental Infanticide". In Wolff, Jerry O.; Sherman, Paul W. (eds.). Rodent Societies: An Ecological and Evolutionary Perspective. הוצאת אוניברסיטת שיקגו. pp. 274–278. ISBN 978-0-226-90538-9.
- ^ Hoogland, J. L. (1985). "Infanticide in prairie dogs: Lactating females kill offspring of close kin". מַדָע. 230 (4729): 1037–1040. ביבקוד:1985Sci...230.1037H. דוי:10.1126/science.230.4729.1037. PMID 17814930. S2CID 23653101.
- ^ Hackländera, Klaus; Möstlb, Erich; Arnold, Walter (2003). "Reproductive suppression in female Alpine marmots, Marmota marmota". התנהגות בעלי חיים. 65 (6): 1133–1140. דוי:10.1006/anbe.2003.2159. S2CID 53218701.
- ^ א ב Charters, Jessie Blount Allen (1904). "The associative processes of the guinea pig: A study of the psychical development of an animal with a nervous system well medullated at birth". Journal of Comparative Neurology and Psychology. XIV (4): 300–337.
- ^ Jacobs, Lucia F.; Liman, Emily R. (1991). "Grey squirrels remember the locations of buried nuts" (PDF). התנהגות בעלי חיים. 41: 103–110. דוי:10.1016/s0003-3472(05)80506-8. S2CID 50448069.
- ^ Jacobs, Lucia F. (1992). "Memory for cache locations in Merriam's kangaroo rats" (PDF). התנהגות בעלי חיים. 43 (4): 585–593. דוי:10.1016/S0003-3472(05)81018-8. S2CID 14173113. הועבר לארכיון מ המקורי (PDF) ב- 26 באוגוסט 2014.
- ^ Harding, E. J.; Paul, E. S.; Mendl, M. (2004). "Animal behaviour: Cognitive bias and affective state". טֶבַע. 427 (6972): 312. ביבקוד:2004Natur.427..312H. דוי:10.1038/427312a. PMID 14737158. S2CID 4411418.
- ^ ריגולה, רפאל; פלוטה, הלנה; פופיק, פיוטר (2012). "חולדות צוחקות אופטימיות". PLOS ONE. 7 (12): e51959. ביבקוד:2012PLoSO ... 751959R. דוי:10.1371 / journal.pone.0051959. PMC 3530570. PMID 23300582.
- ^ קרלייל, קים (8 במרץ 2007). "חולדות המסוגלות לשקף תהליכים נפשיים". אוניברסיטת ג'ורג'יה. הוחזר 13 באוגוסט 2014.
- ^ פוט, אליסון ל. קריסטל, ג'יי ד '(2007). "מטה-קוגניציה בחולדה". ביולוגיה נוכחית. 17 (6): 551–555. דוי:10.1016 / j.cub.2007.01.061. PMC 1861845. PMID 17346969.
- ^ סמית ', ג'יי דייוויד; Beran, M. J .; קושמן, ג'יי ג'יי; Coutinho, M. V. C. (2008). "המחקר ההשוואתי של מטה-קוגניציה: פרדיגמות חדות יותר, מסקנות בטוחות יותר". עלון פסיכונומי וביקורת. 15 (4): 679–691. דוי:10.3758 / PBR.15.4.679. PMC 4607312. PMID 18792496.
- ^ יוזפובייז, י. סטדדון, ג'יי א. ר .; Cerutti, D. T. (2009). "מטה-הכרה בבעלי חיים: איך נדע שהם יודעים?". סקירות הכרה והתנהגות השוואתיות. 4: 29–39. דוי:10.3819 / ccbr.2009.40003.
- ^ הנסון, אן (2012). "איך חולדות בוחרות מה לאכול?". התנהגות חולדות וביולוגיה. הוחזר 24 באוגוסט 2014.
- ^ גאלף, בנט ז '; ללנד, קווין נ '(יוני 2005). "למידה חברתית בבעלי חיים: לימודים אמפיריים ומודלים תיאורטיים". BioScience. 55 (6): 489–499. דוי:10.1641 / 0006-3568 (2005) 055 [0489: sliaes] 2.0.co; 2. JSTOR 10.1641 / 0006-3568% 282005% 29055% 5B0489% 3 ASLIAES% 5D2.0.CO% 3B2.
- ^ א ב ג קיי, אמילי ה '; Hoekstra, Hopi E. (2008). "מכרסמים". ביולוגיה נוכחית. 18 (10): R406 – R410. דוי:10.1016 / j.cub.2008.03.019. PMID 18492466.
- ^ אשר 1, רוברט י. מנג, ג'ין; וויבל, ג'ון ר. מקנה, מלקולם ג. רוג'יר, גילרמו וו .; דשוועג, דמברלין; נובאצ'ק, מייקל ג'יי (2005). "גזע לגומורפה ועתיקות הזוהר". מַדָע. 307 (5712): 1091–1094. ביבקוד:2005Sci ... 307.1091A. דוי:10.1126 / science.1107808. PMID 15718468. S2CID 42090505.
- ^ Douzery, E. J. P .; Delsuc, F .; סטנהופ, מ 'ג'יי; Huchon, D. (2003). "שעונים מולקולריים מקומיים בשלושה גנים גרעיניים: זמני סטייה של מכרסמים ויונקים אחרים ואי התאמה בין כיול מאובנים". כתב העת לאבולוציה מולקולרית. 57: S201–13. ביבקוד:2003JMolE..57S.201D. דוי:10.1007 / s00239-003-0028-x. PMID 15008417. S2CID 23887665.
- ^ הורנר, ד. ס .; לפקימיאטיס, ק '; רייס, א .; גיסי, ג. סאקון, ג. Pesole, G. (2007). "ניתוחים פילוגנטיים של רצפי גנום מיטוכונדריאליים מלאים מצביעים על סטייה בסיסית של המכרסם החידתי. אנומאלורוס". ביולוגיה אבולוציונית של BMC. 7 (1): 16. דוי:10.1186/1471-2148-7-16. PMC 1802082. PMID 17288612.
- ^ ווד, ד 'ג'וזף (2010). הכחדה של הצוללות הרבות מחוץ לצפון אמריקה: גישה עולמית לבדיקת מודל התחרות (גברת.). אוניברסיטת מדינת אוהיו.
- ^ שנק, ג'ון ג'יי; רו, קווין סי .; סטפן, סקוט ג'יי (2013). "הזדמנות אקולוגית ונטייה לגיוון בהתיישבות יבשתית חוזרת ונשנית על ידי מכרסמים מורואידים". ביולוגיה שיטתית. 62 (6): 837–864. דוי:10.1093 / sysbio / syt050. PMID 23925508.
- ^ הופקינס, סמנתה ש.ב. (2005). "האבולוציה של הפוטוריאליות והתפקיד ההסתגלותי של הקרניים במילאגולידה (ממליה: רודנטיה)". הליכים של החברה המלכותית ב. 272 (1573): 1705–1713. דוי:10.1098 / rspb.2005.3171. PMC 1559849. PMID 16087426.
- ^ סמואלס, יהושע X .; זנקנלה, ג'ון (2011). "הופעה מוקדמת של המפיליה של בִּיבָר (Castoridae) מתצורת העכשושים באורגון " (PDF). כתב העת לפליאונטולוגיה. 85 (5): 930–935. דוי:10.1666/11-016.1. S2CID 128866799.
- ^ א ב מריבו, לורן; אסיד, אל מברוק; מרזוגוי, ויסם; עמר, חייט חייאטי; אדנט, סילבן; מרנדט, ברנרד; מרזראוד, ז'יל; טאבוס, רודולף; ויאני-ליאוד, מוניק (2014). "זן חדש ופרימיטיבי של פרוטופיומיות (רודנטיה, Hystricognathi) מהאאוקן האמצעי המאוחר של ג'בל אל קבאר, מרכז תוניסיה ". Palaeovertebrata. 38 (1): 1–17. דוי:10.18563 / pv.38.1.e2.
- ^ גרברנט, עמנואל; זעם, ז'אן קלוד (2006). "פליוביאוגרפיה של אפריקה: עד כמה מובחנת מגונדוואנה ולוראסיה?". פליאוגרפיה, פליאוקלימטולוגיה, פליאולוגיה. 241 (2): 224–246. ביבקוד:2006 PPP ... 241..224 גרם. דוי:10.1016 / j.palaeo.2006.03.016.
- ^ ולז-חורבה, חורחה; מרטין, תומאס; מקפי, רוס ד '(2014). "המכרסמים הקריביים הקדומים ביותר: קוויוממורפים של אוליגוקן מפוארטו ריקו". כתב העת לפליאונטולוגיה של חוליות. 34 (1): 157–163. דוי:10.1080/02724634.2013.789039. S2CID 140178414.
- ^ עלי, ג'יי ר; Huber, M. (20 בינואר 2010). "המגוון הביולוגי של היונקים על מדגסקר הנשלט על ידי זרמי הים". טֶבַע. 463 (4 בפברואר 2010): 653–656. ביבקוד:2010 טבע 463..653 א. דוי:10.1038 / nature08706. PMID 20090678. S2CID 4333977.
- ^ וקואה, א .; בנדוקידזה, או. בוכסיאנידזה, מ '; ואנישווילי, נ '; אוגוסטי, י. מרטינז-נבארו, ב '; רוק, ל '(2010). "דורבן בנאוגן המאוחר וברביעי הרביעי של גרוזיה" (PDF). עלון האקדמיה הלאומית למדעים בגאורגיה. 4 (3): 140–149. הועבר לארכיון מ המקורי (PDF) ב- 16 ביולי 2014.
- ^ "בונה ענק". מחברות להיסטוריה של הטבע. מוזיאון הטבע הקנדי. 28 במאי 2013. הוחזר 19 באוקטובר 2014.
- ^ רינדרנקנכט, אנדרס; בלאנקו, ר 'ארנסטו (2008). "המכרסם המאובן הגדול ביותר". הליכים של החברה המלכותית ב. 275 (1637): 923–928. דוי:10.1098 / rspb.2007.1645. PMC 2599941. PMID 18198140.
- ^ גזע, ביל; פורד, פרד (2007). עכברים עכברושים וחולדות (PDF). הוצאת CSIRO. עמ '3, 5 ופסים. ISBN 978-0-643-09166-5.
- ^ "תוכנית הפעולה למכרסמים אוסטרלים". איכות הסביבה אוסטרליה. 1 באפריל 1995. הוחזר 18 בספטמבר 2014.
- ^ רו, ק. ג. רינו, מ 'ל'; ריצ'מונד, ד 'מ'; אדקינס, ר 'מ'; Steppan, S. J. (2008). "קולוניזציה של פליוקן והקרנות אדפטיביות באוסטרליה ובגינאה החדשה (סאהול): שיטתיות רב-מיקודית של המכרסמים האנדמיים הישנים (Muroidea: Murinae)". פילוגנטיקה מולקולרית ואבולוציה. 47 (1): 84–101. דוי:10.1016 / j.ympev.2008.01.001. PMID 18313945.
- ^ בסקין, ג'ון א .; תומאס, רוני ג'י (2007). "דרום טקסס והמחלף האמריקאי הגדול" (PDF). איגוד עסקאות חברות גיאולוגיות בחוף המפרץ. 57: 37–45. הועבר לארכיון מ המקורי (PDF) ב- 18 ביולי 2014.
- ^ מרשל, ל 'ג'; באטלר, ר. פ .; דרייק, ר 'ה'; קרטיס, ג 'ה'; טדפורד, ר 'ה' (1979). "כיול המחלף האמריקאי הגדול". מַדָע. 204 (4390): 272–279. ביבקוד:1979Sci ... 204..272M. דוי:10.1126 / science.204.4390.272. PMID 17800342. S2CID 8625188.
- ^ א ב סמית ', מרגרט פ. פאטון, ג'יימס ל '(1999). "קשרים פילוגנטיים והקרנת מכרסמים של סיגמודונטין בדרום אמריקה: עדויות מציטוכרום ב". כתב העת להתפתחות היונקים. 6 (2): 89–128. דוי:10.1023 / ת: 1020668004578. S2CID 22355532.
- ^ א ב פאראדה, א .; Pardiñas, U. F. J .; סלזאר-בראבו, י. ד'אליה, ג '; פלמה, ר 'א' (מרץ 2013). "תאריך קרינה ניאו-טרופית מרשימה: הערכות זמן מולקולריות לסיגמודונטינה (רודנטיה) מספקות תובנות לגבי הביוגרפיה ההיסטורית שלה". פילוגנטיקה מולקולרית ואבולוציה. 66 (3): 960–968. דוי:10.1016 / j.ympev.2012.12.001. PMID 23257216.
- ^ סטפן, סקוט ג'יי (18 באפריל 2006). "רודנטיה". פרויקט האינטרנט של עץ החיים. הוחזר 14 ביולי 2014.
- ^ "מכרסם (נ ')". מילון אתיולוגיה מקוון. הוחזר 7 במאי 2015.
- ^ סמית ', אנדרו ט. "לגומורף". אנציקלופדיה בריטניקה. אנציקלופדיה בריטניקה. הוחזר 11 באוגוסט 2014.
- ^ וו, שאויואן; וו, וניו; ג'אנג, פוחון; כן, ג'י; ני, שיג'ון; שמש, ג'ימין; אדוארדס, סקוט החמישי; מנג, ג'ין; אורגן, כריס ל '(2012). "עדויות מולקולריות ופליאונטולוגיות למקור מכרסמים שלאחר הקרטיקון". PLOS ONE. 7 (10): e46445. ביבקוד:2012PLoSO ... 746445W. דוי:10.1371 / journal.pone.0046445. PMC 3465340. PMID 23071573.
- ^ פאברה; ואח '. (2012). "הצצה על תבנית פיזור המכרסמים: גישה פילוגנטית". ביולוגיה אבולוציונית של BMC. 12: 88. דוי:10.1186/1471-2148-12-88. PMC 3532383. PMID 22697210.
- ^ א ב ווד, אלברט א '(1955). "סיווג מתוקן של המכרסמים". כתב העת לממלוגיה. 36 (2): 165–187. דוי:10.2307/1375874. JSTOR 1375874.
- ^ ווד, אלברט א '(1958). "האם יש סדרי משנה למכרסמים?". ביולוגיה שיטתית. 7 (4): 169–173. דוי:10.2307/2411716. JSTOR 2411716.
- ^ קרלטון, מ 'ד'; Musser, G. G. (2005). "הזמנת רודנטיה". בווילסון, דון אי .; רידר, דיאן מ '(עורכים). מינים של יונקים בעולם - התייחסות טקסונומית וגיאוגרפית. 12. לחץ על JHU. עמ '745-752. ISBN 978-0-8018-8221-0.
- ^ Honeycutt, Rodney L. (2009). "מכרסמים (רודנטיה)" (PDF). בגדרות, ס.ב .; קומאר, ש '(עורכים). עץ החיים. הוצאת אוניברסיטת אוקספורד.
- ^ א ב אמורי, ג '; Gippoliti, S. (2003). "גישה גבוהה יותר של סיכון עדיפויות לשימור מכרסמים במאה ה -21". מגוון ביולוגי ושימור בעלי חיים. 26 (2): 1–18.
- ^ Morgan, G. S. (1993). "חוליות יבשה רביעיות של ג'מייקה". ביו-סטרטיגרפיה של ג'מייקה. האגודה הגיאולוגית של אמריקה. 182. עמ '417–442. דוי:10.1130 / mem182-p417. ISBN 978-0-8137-1182-9.
- ^ "הערכת שימור מכרסמים". ווזה. הועבר לארכיון מ המקורי ב -15 ביולי 2014. הוחזר 27 ביוני 2014.
- ^ גודינאס, אדוארדו (1989). לידיקר, ויליאם זנדר (עורך). מכרסמים: סקר עולמי בנושא דאגות לשימור: מבוסס על הליכי סדנה של קבוצת המומחים למכרסמים IUCN / SSC, שהתקיימה בקונגרס הבינלאומי הרביעי, 17 באוגוסט 1985, אדמונטון, אלברטה, קנדה. IUCN. עמ ' 23.
- ^ א ב באקל, א 'פ'; Fenn, M. G. P. (1992). "הדברת מכרסמים בשימור מינים בסכנת הכחדה". ההליכים של ועידת הדברה החולייתנית ה -15: נייר 12. 3–5 במרץ 1992
- ^ הינדווד, ק.א. (1940). "ציפורי האי לורד האו". אֶמוּ. 40: 1–86. דוי:10.1071 / mu940001.
- ^ "Lundy Puffins חזרה מהסף". BBC דבון. 22 בפברואר 2008. הוחזר 30 ביוני 2014.
- ^ מיטשל, הת'ר (27 במאי 2014). "פופינס א-שפע? תקווה חדשה עבור לונדי ואיי עופות-ים אחרים בבריטניה". RSPB. הוחזר 30 ביוני 2014.
- ^ פיינשטיין, קלי (1 במרץ 2006). "להגיש כובע בונה". פרווה לבד אופנתית. UC סנטה קרוז. הועבר לארכיון מ המקורי ב- 6 באוקטובר 2014. הוחזר 24 בספטמבר 2014.
- ^ איניס, הרולד א. (1999). סחר הפרווה בקנדה: מבוא להיסטוריה הכלכלית הקנדית. הוצאת אוניברסיטת טורונטו. עמ '9–12. ISBN 978-0-8020-8196-4.
- ^ א ב "סחר מוגזם: בגדים וחיתוך". גן החיות של גרנבי. מאי 2010. הוחזר 9 באוגוסט 2014.
- ^ "אמנות ואגדה של Quillwork של Lakota". מוזיאון אקטה לקוטה ומרכז תרבות. הוחזר 29 ביוני 2013.
- ^ א ב ג ד פידלר, לינווד א '(1990). "מכרסמים כמקור מזון". ההליכים של ועידת הדברה החולייתית הארבע-עשרה 1990: 149–155.
- ^ נואלטון, דיוויד (13 ביולי 2011). "שפן ניסיונות, ארוחה מחמדת או חגיגית". קוזקו אוכל. הועבר לארכיון מ המקורי ב- 14 ביולי 2014. הוחזר 5 ביולי 2014.
- ^ מורוולי, וולטר קוראזה; נואלטון, דיוויד (5 במרץ 2012). "תנורי בוץ ותנורים מסורתיים מכינים את האוכל הטוב ביותר". קוזקו אוכל. הועבר לארכיון מ המקורי ב -15 ביולי 2014. הוחזר 6 ביולי 2014.
- ^ וולף, ג'רי או. שרמן, פול וו. (2008). אגודות מכרסמים: נקודת מבט אקולוגית ואבולוציונית. הוצאת אוניברסיטת שיקגו. עמ '3–8. ISBN 978-0-226-90538-9.
- ^ קרינקה, ג'ורג 'י. בולוק, גיליאן ר.; בונטון, טרייסי (2000). "היסטוריה, זנים ודגמים". חולדת המעבדה (מדריך לחיות ניסוי). העיתונות האקדמית. עמ '3–16. ISBN 978-0-12-426400-7.
- ^ מורס, הרברט סי (1981). "עכבר המעבדה: הערכה היסטורית". בפוסטר, הנרי (עורך). העכבר במחקר ביו-רפואי: היסטוריה, גנטיקה ועכברים פראיים. אלסבייה. עמ.xi, 1. ISBN 978-0-323-15606-6.
- ^ א ב גד, שיין סי (2007). מודלים של בעלי חיים בטוקסיקולוגיה (מהדורה שנייה). טיילור ופרנסיס. עמ '334-402. ISBN 978-0-8247-5407-5.
- ^ הרקנס, ג'ון א. וגנר, ג'וזף א '(1995). הביולוגיה והרפואה של ארנבים ומכרסמים. וויליאמס ווילקינס. עמ '30–39. ISBN 978-0-683-03919-1.
- ^ גרריני, אניטה (2003). התנסות בבני אדם ובעלי חיים. ג'ונס הופקינס. עמ '98-104. ISBN 978-0-8018-7196-2.
- ^ גריי, טארה (1998). "היסטוריה קצרה של בעלי חיים בחלל". מנהל האווירונאוטיקה והחלל הלאומי. הוחזר 5 במרץ 2007.
- ^ שרווין, סי מ '(2010). "25: גידול ורווחת מכרסמים לא מסורתיים במעבדה". בהוברכט, ר '; קירקווד, ג'יי (עורכים). מדריך UFAW בנושא טיפול וניהול של חיות מעבדה. וויילי-בלקוול. עמ '359–369.
- ^ יינות, מייקל (19 במאי 2004). "מכרסמים בגמביה מסתכנים במוות בגלל בננות". הגיל. הוחזר 21 ביוני 2014.
- ^ מהללה, חסן (13 בספטמבר 2012). "חולדות ענק שהוכשרו לגילוי מוקשים ושחפת באפריקה". BBC. הוחזר 27 ביוני 2014.
- ^ בקאלאר, ניקולס (3 בינואר 2011). "גילוי שחפת: אין מיקרוסקופים, רק חולדות". ניו יורק טיימס. הוחזר 23 באוגוסט 2014.
- ^ Harder, Ben (1 במאי 2002). "מדענים" מניעים "חולדות באמצעות שלט רחוק". נשיונל גאוגרפיק. הוחזר 9 בנובמבר 2013.
- ^ סולון, או '(9 בספטמבר 2013). "המשימה של האדם לבנות מערכות שליטה מרחוק לכלבים, מקקים וכרישים". קווית. הועבר לארכיון מ המקורי ב -4 בנובמבר 2013. הוחזר 9 בדצמבר 2013.
- ^ שו, ש .; טלוואר, ס. ק .; Hawley, E. S .; לי, ל '; Chapin, J. K. (2004). "מערכת טלמטריה רב-ערוצית לגירוי מיקרו-מוחי בבעלי חיים משוטטים בחופשיות". כתב העת לשיטות מדעי המוח. 133 (1–2): 57–63. דוי:10.1016 / j.jneumeth.2003.09.012. PMID 14757345. S2CID 10823.
- ^ "שרקנים". RSPCA. 2014. הוחזר 21 ביוני 2014.
- ^ "מכרסמים לחיות מחמד". RSPCA. 2014. הוחזר 21 ביוני 2014.
- ^ ברוקל, ריי (1983). חיות מחמד של גרביל ומכרסמים קטנים אחרים. עיתונות ילדים. עמ '5–20. ISBN 978-0-516-01679-5.
- ^ מירבורג, ב 'ג'; סינגלטון, ג 'ר; Leirs, H. (2009). "שנת החולדה מסתיימת: הזמן להילחם ברעב!". מדעי ניהול מזיקים. 65 (4): 351–2. דוי:10.1002 / ps.1718. PMID 19206089.
- ^ א ב סטנשת, נילס כר; לירס, הרוויג; סקונהופט, אנדרס; דייויס, סטיבן א .; פץ ', רוג'ר פ. אנדריסן, הארי פ .; סינגלטון, גרנט ר .; לימה, מאוריסיו; מצ'אנג'ו, רוברט ס .; מקונדי, רודוס ה '; ג'אנג, ג'יבין; בראון, פיטר ר. שי, דז'או; וואן, שינרונג (2003). "עכברים, חולדות ואנשים: הביו-כלכלה של מזיקים למכרסמים חקלאיים". גבולות באקולוגיה ובסביבה. 1 (77): 367–375. דוי:10.2307/3868189. JSTOR 3868189.
- ^ א ב מירבורג, ב 'ג'; סינגלטון, ג 'ר'; קילסטרה, א '(2009). "מחלות המועברות במכרסמים והסיכונים שלהן לבריאות הציבור". ביקורות קריטיות במיקרוביולוגיה. 35 (3): 221–70. דוי:10.1080/10408410902989837. PMID 19548807. S2CID 205694138.
- ^ McCormick, M. (2003). "חולדות, תקשורת ומגפה: לקראת היסטוריה אקולוגית" (PDF). כתב העת להיסטוריה בינתחומית. 34 (1): 1–25. דוי:10.1162/002219503322645439. S2CID 128567627. הועבר לארכיון מ המקורי (PDF) ב- 22 בפברואר 2015.
- ^ "מחלות המועברות במכרסמים". המרכז האירופי למניעת ובקרת מחלות. הועבר לארכיון מ המקורי ב -3 בספטמבר 2014. הוחזר 1 בספטמבר 2014.
- ^ "מחלות המועברות בעקיפין על ידי מכרסמים". מרכזים לבקרת מחלות ומניעתן. 2012. הוחזר 1 בספטמבר 2014.
- ^ מרכזים לבקרת מחלות ומניעתן (2006). ניהול מזיקים משולב: עריכת סקרי מכרסמים עירוניים (PDF). אטלנטה: משרד הבריאות והשירותים האנושיים בארה"ב.
- ^ Wodzicki, K. (1973). "סיכויים לשליטה ביולוגית באוכלוסיות מכרסמים". עלון ארגון הבריאות העולמי. 48 (4): 461–467. PMC 2481104. PMID 4587482.
- ^ "הדברת מכרסמים בחקלאות - מדריך HGCA". המועצה לפיתוח חקלאות וגננות. 2012. הוחזר 24 בפברואר 2018.
לקריאה נוספת
- מקנה, מלקולם ג. בל, סוזן ק. (1997). סיווג יונקים מעל רמת המינים. הוצאת אוניברסיטת קולומביה. ISBN 978-0-231-11013-6.
- ווילסון, ד 'א'; רידר, ד. מ., עורכים. (2005). מינים של יונקים בעולם: התייחסות טקסונומית וגיאוגרפית. הוצאת אוניברסיטת ג'ונס הופקינס. ISBN 978-0-8018-8221-0.
- קרלטון, מ 'ד'; Musser, G. G. "Order Rodentia", עמודים 745–752 בווילסון ורידר (2005).
קישורים חיצוניים
זואולוגיה, אוסטולוגיה, אנטומיה השוואתית
שׁוֹנִים
- מכרסמים אפריקאים
- תמונות מכרסמים
- דפי עובדות על סוגים של מכרסמים מ ה האגודה הארצית להדברת מזיקים על עכברי צבאים, חולדות נורבגיה ומיני מכרסמים אחרים