| מערכת נשימה | |
|---|---|
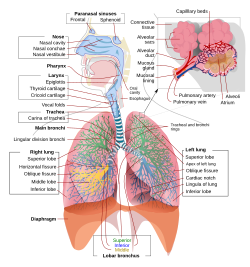 מבט סכמטי שלם על מערכת הנשימה האנושית על חלקיה ותפקידיה. | |
| פרטים | |
| מזהים | |
| לָטִינִית | מערכת הנשימה מערכתית |
| רֶשֶׁת | D012137 |
| ת"א 98 | A06.0.00.000 |
| TA2 | 3133 |
| FMA | 7158 |
| מינוח אנטומי | |
ה מערכת נשימה (גַם מכשירי נשימה, מערכת אוורור) הוא מערכת ביולוגית המורכב מספציפי איברים ומבנים המשמשים ל חילופי גז ב חיות ו צמחים. האנטומיה והפיזיולוגיה שגורמים לכך לקרות משתנים מאוד, תלוי בגודל האורגניזם, בסביבה בה הוא חי ובהיסטוריה האבולוציונית שלו. ב חיות יבשה משטח הנשימה מופנם כציפוי של הנשימה ריאות.[1] חילופי גז בריאות מופיע במיליוני שקיות אוויר קטנות הנקראות alveoli אצל יונקים וזוחלים, אבל אטריה אצל ציפורים. לשקי האוויר המיקרוסקופיים הללו אספקת דם עשירה מאוד, ובכך מביאים את האוויר למגע קרוב עם הדם.[2] שקיות אוויר אלה מתקשרות עם הסביבה החיצונית באמצעות מערכת דרכי הנשימה, או צינורות חלולים, שהגדול ביותר הוא קנה הנשימה, שמסתעף באמצע החזה לשני העיקריים סימפונות. אלה נכנסים לריאות שם הם מסתעפים לסימפונות משניים ושלישוניים הצרים בהדרגה שמתפצלים במספר צינורות קטנים יותר ברונכיולות. ב ציפורים הסימפונות נקראות פרברונצ'י. זה הסימפונות, או הפרברונצ'י שנפתחים בדרך כלל לתוך המיקרוסקופי alveoli אצל יונקים ו אטריה אצל ציפורים. יש לשאוב אוויר מהסביבה אל הכליות או אטריה בתהליך של נְשִׁימָה הכוללת את שרירי נשימה.
ברוב דג, ומספר אחרים בעלי חיים ימיים (שניהם בעלי חוליות ו חסרי חוליות ) מערכת הנשימה מורכבת מ זימים, שהם איברים חיצוניים חלקית או מלאה, שטופים בסביבה המימית. מים אלה זורמים מעל הזימים במגוון אמצעים פעילים או פסיביים. החלפת גז מתרחשת בזימים המורכבים מדקים או שטוחים מאוד חוטים ו ממלי שחושפים שטח פנים גדול מאוד של כלי דם רקמות למים.
בעלי חיים אחרים, כגון חרקים, יש מערכות נשימה עם מאפיינים אנטומיים פשוטים מאוד, ובפנים דו-חיים אפילו ה עור ממלא תפקיד חיוני בהחלפת הגז. צמחים יש גם מערכות נשימה, אך כיוון החלפת הגז יכול להיות מנוגד לבעלי חיים. מערכת הנשימה בצמחים כוללת מאפיינים אנטומיים כגון סטומטה, הנמצאים בחלקים שונים של הצמח.[3]
יונקים
אֲנָטוֹמִיָה

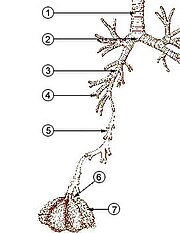
ב בני אנוש ואחר יונקים האנטומיה של מערכת נשימה טיפוסית היא דרכי הנשימה. המסכת מחולקת ל- עֶלִיוֹן ו דרכי הנשימה התחתונות. הדרך העליונה כוללת את אף, חללים באף, סינוסים, לוֹעַ ואת החלק של גָרוֹן מעל ה קפלי קול. הדרך התחתונה (איור 2.) כוללת את החלק התחתון של הגרון, את קנה הנשימה, סימפונות, ברונכיולות וה alveoli.
דרכי הנשימה המסועפות של דרכי התחתון מתוארות לעתים קרובות כ- עץ נשימה או עץ טרכאו-ברונכיאלי (איור 2).[4] המרווחים בין נקודות הסתעפות עוקבות לאורך הענפים השונים של "העץ" מכונים לעתים קרובות "דורות" מסועפים, אשר אצל האדם הבוגר בסביבות 23. הדורות הקודמים (בערך הדורות 0–16), המורכבים מ קנה הנשימה והסמפונות, כמו גם הסמפונות הגדולות יותר הפועלות כמו תעלות אוויר, מביא אוויר לברונכיוליות הנשימה, צינורות המכתש והכליות (בערך הדורות 17-23), שם חילופי גז מתרחש.[5][6] ברונכיולות מוגדרים כדרכי הנשימה הקטנות חסרות כל תמיכה סחוסית.[4]
הסמפונות הראשונות שהסתעפו מה- קנה הנשימה הם הסמפונות העיקריים מימין ומשמאל. שנייה רק בקוטר לקנה הנשימה (1.8 ס"מ), הסמפונות הללו (בקוטר -1 -1.4 ס"מ)[5] להיכנס ל ריאות בכל הילום, שם הם מסתעפים לסמפונות משניים צרים יותר המכונים סימפונות לובר, ואלה מסתעפים לסמפונות שלישוניים צרים יותר המכונים סימפונות קטעיות. חלוקות נוספות של הסמפונות הסגמנטליים (בקוטר 1 עד 6 מ"מ)[7] ידועים כסימפונות פלחים מסדר רביעי, מסדר 5 ובסדר 6, או מקובצים יחד כסמפונות תת-מגזריים.[8][9]
בהשוואה למספר 23 (בממוצע) של הסתעפות עץ הנשימה אצל האדם הבוגר, עכבר יש רק כ- 13 הסתעפות כזו.
המכתשים הם מסופי מבוי סתום של "העץ", כלומר כל אוויר שנכנס אליהם צריך לצאת באותו מסלול. מערכת כזו יוצרת שטח מת, נפח אוויר (כ -150 מ"ל בבני האדם הבוגר) הממלא את דרכי הנשימה לאחר הנשיפה ומונשם חזרה אל הכליות לפני שהאוויר הסביבתי מגיע אליהם.[10][11] בתום השאיפה דרכי הנשימה מלאות באוויר סביבתי שננשף מבלי לבוא במגע עם מחליף הגז.[10]
נפחי אוורור
הריאות מתרחבות ומתכווצות במהלך מחזור הנשימה, תוך שאיבת אוויר לריאות ומחוצה לה. נפח האוויר נע או החוצה מהריאות בנסיבות מנוחה רגילות (המנוחה נפח גאות ושפל כ 500 מ"ל), ונפחים הנעים במהלך שאיפה מאולצת מקסימאלית ונשיפה מאולצת מקסימאלית נמדדים בבני אדם על ידי ספירומטריה.[12] ספירוגרמה אנושית מבוגרת טיפוסית עם השמות שניתנו לטיולים השונים בנפח שהריאות יכולות להופיע מודגמת להלן (איור 3):

לא כל האוויר בריאות יכול להיפלט במהלך נשיפה מאולצת. זה נפח שיורי של כ 1.0-1.5 ליטר שלא ניתן למדוד באמצעות ספירומטריה. נפחים הכוללים את הנפח השיורי (כלומר יכולת שיורית פונקציונלית של כ 2.5-3.0 ליטר, ו קיבולת ריאות כוללת ולכן לא ניתן למדוד אותם באמצעות ספירומטריה. המדידה שלהם דורשת טכניקות מיוחדות.[12]
קצב הנשימה של האוויר פנימה או החוצה, דרך הפה או האף, או אל תוך כריתה של הכליות מתואר בטבלה הבאה, יחד עם אופן חישובם. מספר מחזורי הנשימה לדקה מכונה קצב נשימה.
| מדידה | משוואה | תיאור |
|---|---|---|
| אוורור דקה | נפח גאות ושפל * קצב נשימה | נפח האוויר הכולל שנכנס, או עוזב, את האף או את הפה בדקה. |
| אוורור מכתש | (נפח גאות ושפל - שטח מת) * קצב נשימה | נפח האוויר שנכנס או יוצא מהאלבולות לדקה. |
| אוורור חלל מת | שטח מת * קצב נשימה | נפח האוויר שלא מגיע לאלבולי במהלך שאיפה, אלא נשאר בדרכי הנשימה, לדקה. |
מכניקות נשימה



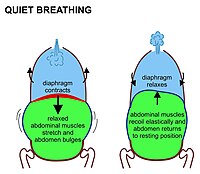

ב יונקים שאיפה במנוחה נובעת בעיקר מהתכווצות דִיאָפרַגמָה. זהו סדין שרירים כיפה כלפי מעלה המפריד בין חלל בית החזה לחלל הבטן. כאשר הוא מתכווץ הסדין משתטח, (כלומר נע כלפי מטה כפי שמוצג באיור 7) ומגדיל את נפח חלל בית החזה. הסרעפת המתכווצת דוחפת את אברי הבטן כלפי מטה. אך מכיוון שרצפת האגן מונעת את איברי הבטן התחתונה ביותר לנוע בכיוון זה, תוכן הבטן הגמיש גורם לבטן להתנפח כלפי חוץ לפנים ולצדדים, מכיוון ששרירי הבטן הנינוחים אינם מתנגדים לתנועה זו (איור 7). בליטה פסיבית לחלוטין (ומתכווצת בזמן נשיפה) של הבטן בזמן נשימה רגילה מכונה לעיתים "נשימת בטן", אם כי למעשה מדובר ב"נשימה סרעפתית ", שאינה נראית מבחוץ בגוף. יונקים משתמשים בשרירי הבטן שלהם רק בזמן נשיפה חזקה (ראה איור 8 והדיון בהמשך). אף פעם לא בשאיפה כלשהי.
כאשר הסרעפת מתכווצת, בית החזה מוגדל בו זמנית על ידי הצלעות הנמשכות כלפי מעלה על ידי שרירי צלעי כפי שמוצג באיור 4. כל הצלעות משופעות כלפי מטה מאחור לחזית (כפי שמוצג באיור 4); אבל הצלעות התחתונות גַם משופע כלפי מטה מקו האמצע כלפי חוץ (איור 5). כך ניתן להגדיל את הקוטר הרוחבי של כלוב הצלעות באותה צורה בה גדל הקוטר האנטר-אחורי על ידי מה שמכונה תנועת ידית משאבה מוצג באיור 4.
הגדלת המימד האנכי של חלל בית החזה על ידי התכווצות הסרעפת, ושני המימדים האופקיים שלה באמצעות הרמת חזית וצלעות הצלעות, גורמים ליפול הלחץ התוך-חזי. פנים הריאות פתוחים לאוויר החיצוני, והיותם אלסטיים, ולכן מתרחבים למלא את החלל המוגדל. הזרמת האוויר לריאות מתרחשת דרך דרכי הנשימה בדרכי הנשימה (איור 2). בבריאות, דרכי הנשימה האלה התחל עם האף.[13][14] אפשר להתחיל עם הפה, שהיא מערכת נשימת הגיבוי. עם זאת, כרוני נשימה בפה מוביל למחלה, או שהוא סימן למחלה.[15][16][17] הם מסתיימים בשקיות ללא מוצא מיקרוסקופיות שנקראות alveoli ) פתוחים תמיד, אם כי ניתן לשנות את הקוטר של החלקים השונים באמצעות אוֹהֵד ו מערכות עצבים פרה-סימפטיות. לכן לחץ האוויר המכתשי קרוב תמיד ללחץ האוויר האטמוספרי (בערך 100kPa בגובה פני הים) בזמן מנוחה, כאשר שיפועי הלחץ הגורמים לאוויר לנוע פנימה והחוצה מהריאות במהלך הנשימה לעיתים נדירות עולים על 2-3 kPa.[18][19]
בזמן הנשיפה הסרעפת והשרירים הבין-צלעיים נרגעים. זה מחזיר את החזה והבטן למצב שנקבע על פי גמישותם האנטומית. זהו "המיקום האמצעי המנוח" של בית החזה והבטן (איור 7) כאשר הריאות מכילות את שלהן יכולת שיורית פונקציונלית אוויר (האזור הכחול בהיר באיור הימני של איור 7), אשר אצל האדם הבוגר נפחו של כ 2.5-3.0 ליטר (איור 3).[6] נשיפת מנוחה נמשכת כפי שניים משאיפה מכיוון שהסרעפת נרגעת בצורה פסיבית בעדינות רבה יותר מכפי שהיא מתכווצת באופן פעיל במהלך השאיפה.

נפח האוויר שעובר פנימה אוֹ החוצה (באף או בפה) במהלך מחזור נשימה יחיד נקרא נפח גאות ושפל. אצל אדם בוגר במנוחה מדובר בכ -500 מ"ל לנשימה. בתום הנשיפה דרכי הנשימה מכילות כ -150 מ"ל אוויר מכתשי המהווה את האוויר הראשון שנשאף חזרה אל הכליות במהלך השאיפה.[10][20] אוויר נפח זה שנשאף אל הכבד וחוזר פנימה מכונה שטח מת אוורור, שיש לו את התוצאה של 500 מ"ל שנשאף אל הכליות בכל נשימה רק 350 מ"ל (500 מ"ל - 150 מ"ל = 350 מ"ל) הוא אוויר חם ולח רענן.[6] מכיוון ש -350 מ"ל זה של אוויר צח מעורבב ומדולל ביסודיות באוויר שנשאר בכליות לאחר הנשיפה הרגילה (כלומר יכולת שיורית פונקציונלית 2.5-3.0 ליטר), ברור כי הרכב האוויר המכתשי משתנה מעט מאוד במהלך מחזור הנשימה (ראה איור 9). החמצן מתח (או לחץ חלקי) נותר קרוב ל 13-14 kPa (כ 100 מ"מ כספית), וזה של פחמן דו חמצני קרוב מאוד ל 5.3 kPa (או 40 מ"מ כספית). זה מנוגד להרכב האוויר החיצוני היבש בגובה פני הים, כאשר הלחץ החלקי של החמצן הוא 21 קפ"א (או 160 מ"מ כספית) וזה של פחמן דו חמצני 0.04 קפ"א (או 0.3 מ"מ כספית).[6]
בזמן נשימה כבדה (היפרפנאה ), כמו, למשל, במהלך פעילות גופנית, שאיפה נגרמת על ידי טיול חזק יותר וגדול יותר של הסרעפת המתכווצת מאשר במנוחה (איור 8). בנוסף "שרירי שאיפה אביזרים "מגזים בפעולות של השרירים הבין-צלעיים (איור 8). שרירי שאיפה נלווים אלה הם שרירים הנמשכים מה חוליות צוואר הרחם ובסיס הגולגולת לצלעות העליונות ו סטרנום, לפעמים דרך קשר מתווך ל עצמות הבריח.[6] כאשר הם מתכווצים נפח הפנימי של כלוב הצלעות גדל במידה רבה בהרבה ממה שניתן להשיג על ידי כיווץ השרירים הבין-צלעיים בלבד. נראה מחוץ לגוף לעיתים נקראת הרמת עצם הבריח בזמן שאיפה מאומצת או מאומצת נשימה עצם הבריח, נראה במיוחד במהלך אַסְתְמָה התקפות ובאנשים עם מחלת ריאות חסימתית כרונית.
בזמן נשימה כבדה הנשיפה נגרמת על ידי הרפיה של כל שרירי השאיפה. אבל עכשיו, שרירי הבטן במקום להישאר רגועים (כמו במנוחה), מתכווצים בכוח ומושכים את הקצוות התחתונים של בית החזה כלפי מטה (קדמי וצדדים) (איור 8). זה לא רק מקטין באופן דרסטי את גודל כלוב הצלעות, אלא גם דוחף את אברי הבטן כלפי מעלה אל הסרעפת, וכתוצאה מכך בולט עמוק לתוך בית החזה (איור 8). נפח הריאות בסוף הנשיפה נמצא כעת הרבה מתחת למצב האמצעי במנוחה ומכיל הרבה פחות אוויר מ"יכולת השיורית התפקודית "המנוחה. עם זאת, אצל יונק רגיל לא ניתן לרוקן את הריאות לחלוטין. אצל אדם בוגר תמיד נותר לפחות 1 ליטר אוויר שאריות בריאות לאחר הנשיפה המרבית.[6]
הנשימה הקצבית האוטומטית פנימה והחוצה, יכולה להיות מופרעת על ידי שיעול, עיטוש (צורות של נשיפה חזקה מאוד), על ידי הבעת מגוון רחב של רגשות (צחוק, אנחה, בכי בכאב, נזילות נרגשות) ועל ידי כאלה מעשים מרצון כדיבור, שירה, שריקה ונגינת כלי נשיפה. כל הפעולות הללו נשענות על השרירים שתוארו לעיל, והשפעותיהם על תנועת האוויר לריאות ומחוצה לה.
אמנם לא סוג של נשימה, אבל תמרון וולסאלבה כרוך בשרירי הנשימה. זהו, למעשה, מאמץ נשיפה חזק מאוד נגד סגור היטב גלוטי, כך שאף אוויר לא יכול לברוח מהריאות.[21] במקום תוכן הבטן מתפנה בכיוון ההפוך, דרך פתחים ברצפת האגן. שרירי הבטן מתכווצים בעוצמה רבה וגורמים ללחץ בתוך הבטן ובית החזה לעלות לרמות גבוהות במיוחד. התמרון של Valsalva יכול להתבצע באופן וולונטרי, אך בדרך כלל הוא רפלקס שנגרם כשמנסים לרוקן את הבטן בזמן למשל עשיית צרכים קשה או במהלך הלידה. הנשימה מפסיקה במהלך התמרון הזה.
חילופי גז
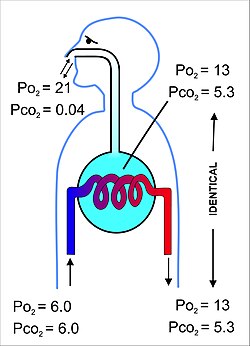


המטרה העיקרית של מערכת הנשימה היא שיווי משקל של הלחצים החלקיים של גזי הנשימה באוויר המכתשי עם אלו הנמצאים בדם הנימי הריאתי (איור 11). תהליך זה מתרחש על ידי פשוט ריכוך,[22] על פני קרום דק מאוד (המכונה מחסום דם – אוויר ), המהווה את קירות ה כבד ריאתי (איור 10). זה מורכב מ תאי אפיתל מכתשי, שלהם קרומי מרתף וה תאי אנדותל של נימי המכתש (איור 10).[23] מחסום גזי הדם הזה הוא דק במיוחד (בבני אדם בעובי 2.2 מיקרומטר בממוצע). הוא מקופל לכ -300 מיליון שקיות אוויר קטנות הנקראות alveoli[23] (כל אחד בקוטר 75 עד 300 מיקרומטר) מסתעף מהנשימה ברונכיולות בתוך ה ריאות ובכך מספק שטח פנים גדול במיוחד (כ -145 מ '2) כדי שחילופי גז יתרחשו.[23]
האוויר הכלול בתוך הכליות מכיל נפח קבוע למחצה של כ- 2.5-3.0 ליטר המקיף לחלוטין את הדם הנימי המכתשי (איור 12). זה מבטיח שיווי משקל של הלחצים החלקיים של הגזים בשני התאים יעיל מאוד ומתרחש במהירות רבה. לדם שעוזב את נימי המכתש ובסופו של דבר מופץ בגוף יש לו לחץ חלקי של חמצן של 13-14 kPa (100 מ"מ כספית), ו- a לחץ חלקי של פחמן דו חמצני של 5.3 kPa (40 מ"מ כספית) (כלומר זהה למתיחות גז החמצן והפחמן הדו-חמצני כמו בכליות).[6] כאמור ב החלק לעיל, הלחצים החלקיים המקבילים של חמצן ופחמן דו חמצני באוויר הסביבתי (היבש) בגובה פני הים הם 21 kPa (160 mmHg) ו- 0.04 kPa (0.3 mmHg) בהתאמה.[6]
ניתן לשמור על ההבדל הניכר בין הרכב האוויר המכתשי לזה של האוויר הסביבתי מכיוון שה- יכולת שיורית פונקציונלית כלול בשקיות ללא מוצא המחוברות לאוויר החיצוני על ידי צינורות צרים וארוכים יחסית (דרכי הנשימה: אף, לוֹעַ, גָרוֹן, קנה הנשימה, סימפונות וענפיהם עד ברונכיולות ), דרכו יש לנשום את האוויר פנימה והחוצה (כלומר אין זרימת דרך חד כיוונית כמו שיש ריאת ציפורים ). אנטומיה טיפוסית זו של יונקים בשילוב העובדה שהריאות אינן מתרוקנות ונפחות מחדש בכל נשימה (משאיר נפח אוויר משמעותי, של כ- 2.5-3.0 ליטר, במכתשים לאחר הנשיפה), מבטיח כי הרכב המכתש. האוויר מופרע רק במינימום כאשר מערבבים אליו את 350 מ"ל האוויר הצח עם כל שאיפה. לפיכך חיה מספקת "אווירה ניידת" מיוחדת מאוד, שהרכבה שונה באופן משמעותי מה אוויר הסביבה של ימינו.[24] זו האווירה הניידת הזו ( יכולת שיורית פונקציונלית ) אליהם נחשף הדם ולכן רקמות הגוף - לא לאוויר החיצוני.
הלחץ החלקי העורקי וכתוצאה מכך של חמצן ופחמן דו חמצני הם מבוקר הומאוסטטי. עלייה בלחץ החלקי העורקי של CO2 ובמידה פחותה, נפילה בלחץ החלקי העורקי של O2, יגרום באופן רפלקסי לנשימה עמוקה ומהירה יותר עד ל מתחים בגזים בדם בריאות, ולכן הדם העורקי, חוזר לקדמותו. ההיפך קורה כאשר מתח הפחמן הדו-חמצני יורד, או, שוב במידה פחותה, מתח החמצן עולה: קצב ועומק הנשימה מופחת עד להחזרת תקינות גזי הדם.
מכיוון שלדם המגיע לנימים המכתשיים יש לחץ חלקי של O2 של, בממוצע, 6 kPa (45 מ"מ כספית), בעוד שהלחץ באוויר המכתשי הוא 13-14 kPa (100 מ"מ כספית), יהיה פיזור נקי של חמצן לדם הנימי, וישנה את הרכב 3 ליטרים אוויר מכתש מעט. באופן דומה, מכיוון שלדם המגיע לנימים המכתשיים יש לחץ חלקי של CO2 של כ- 6 kPa (45 מ"מ כספית), ואילו זה של האוויר המכתשי הוא 5.3 kPa (40 מ"מ כספית), קיימת תנועה נטו של פחמן דו חמצני אל מחוץ לנימים אל הכליות. השינויים שחוללו זרמים נטו אלה של גזים בודדים אל תוך האוויר המכתשי ומחוצה לו מחייבים החלפה של כ -15% מהאוויר המכתשי באוויר הסביבתי כל 5 שניות לערך. זה נשלט בצורה הדוקה מאוד על ידי ניטור גזי הדם העורקיים (המשקפים במדויק את הרכב האוויר המכתשי) על ידי אבי העורקים ו גופות ראש, כמו גם על ידי חיישן גז דם ו- pH על המשטח הקדמי של medulla oblongata במוח. ישנם גם חיישני חמצן ופחמן דו חמצני בריאות, אך הם קובעים בעיקר את הקוטר של ברונכיולות ו נימי ריאות ולכן הם אחראים להפניית זרימת האוויר והדם לחלקים שונים של הריאות.
רק כתוצאה משמירה מדויקת על הרכב 3 ליטר האוויר המכתשי, עם כל נשימה מוזרם קצת פחמן דו חמצני לאטמוספרה וחמצן נלקח מהאוויר החיצוני. אם יותר פחמן דו חמצני מהרגיל אבד בתקופה קצרה של היפרוונטילציה, הנשימה תואט או תיעצר עד שהלחץ החלקי המכתשי של פחמן דו חמצני יחזור ל 5.3 kPa (40 מ"מ כספית). לפיכך, אין זה נכון כי תפקידה העיקרי של מערכת הנשימה הוא להיפטר מגוף מ"פסולת ". את הפחמן הדו-חמצני שמונשם בכל נשימה אפשר כנראה לראות בצורה נכונה יותר כתוצר לוואי של הנוזל החוץ-תאי של הגוף. פחמן דו חמצני ו הומאוסטטים pH
אם הומאוסטטים אלה נפגעים, אז חומצה בדרכי הנשימה, או א אלקלוזיס נשימתי יקרה. בטווח הארוך ניתן לפצות את אלה באמצעות התאמות כליות למערכת ה+ ו- HCO3− ריכוזים בפלזמה; אך מכיוון שזה לוקח זמן, תסמונת היפרוונטילציה יכול, למשל, להתרחש כאשר תסיסה או חרדה גורמים לאדם לנשום מהר ועמוק ובכך לגרום למצוקה אלקלוזיס נשימתי דרך פיצוץ של יותר מדי CO2 מהדם לאוויר החיצוני.[25]
לחמצן מסיסות נמוכה מאוד במים, ולכן הוא נישא בדם בשילוב רופף עם הֵמוֹגלוֹבִּין. החמצן מוחזק על המוגלובין על ידי ארבעה ברזל ברזל -מֵכִיל heme קבוצות לכל מולקולת המוגלובין. כשכל קבוצות ההם נושאות O אחד2 מולקולה שלכל אחת מהדם נאמר שהיא "רוויה" בחמצן, ואף עלייה נוספת בלחץ החלקי של החמצן לא תגדיל משמעותית את ריכוז החמצן של הדם. רוב הפחמן הדו-חמצני בדם מובל כיוני ביקרבונט (HCO3−) בפלזמה. עם זאת ההמרה של CO מומס2 לתוך HCO3− (באמצעות תוספת מים) איטי מדי לקצב שבו הדם מסתובב דרך הרקמות מחד גיסא, ובאמצעות נימים מכתשיים מאידך גיסא. התגובה מזורזת על ידי אנהידראז פחמני, an אֶנזִים בתוך ה תאי דם אדומים.[26] התגובה יכולה ללכת לשני הכיוונים בהתאם ללחץ החלקי הרווח של CO2.[6] כמות קטנה של פחמן דו חמצני מועברת על חלק החלבון של מולקולות ההמוגלובין כ- קרבמינו קבוצות. הריכוז הכולל של פחמן דו חמצני (בצורה של יוני ביקרבונט, CO מומס2, וקבוצות קרבמינינו) בדם עורקי (כלומר לאחר שהוא מאוזן עם האוויר המכתשי) הוא כ 26 מ"מ (או 58 מ"ל / 100 מ"ל),[27] בהשוואה לריכוז החמצן בדם עורקי רווי של כ- 9 מ"מ (או 20 מ"ל / 100 מ"ל דם).[6]
בקרת אוורור
אוורור הריאות ביונקים מתרחש דרך מרכזי נשימה בתוך ה medulla oblongata וה פונס של ה גזע המוח.[6] אזורים אלה מהווים סדרה של מסלולים עצביים שמקבלים מידע על לחצים חלקיים של חמצן ופחמן דו חמצני בתוך ה דם עורקי. מידע זה קובע את קצב האוורור הממוצע של alveoli של ה ריאות, לשמור על אלה לחצים קבועים. מרכז הנשימה עושה זאת באמצעות עצבים מוטוריים אשר מפעילים את דִיאָפרַגמָה ואחר שרירי נשימה.
קצב הנשימה עולה כאשר ה- לחץ חלקי של פחמן דו חמצני בדם עולה. זה מזוהה על ידי כימותרפטי גז בדם מרכזיים על המשטח הקדמי של medulla oblongata.[6] ה אבי העורקים ו גופות ראש, הם כימותרפטי גז בדם היקפיים הרגישים במיוחד לעורק לחץ חלקי של O2 אם כי הם גם מגיבים, אך פחות חזק, ללחץ החלקי של שיתוף2.[6] בגובה פני הים, בנסיבות רגילות, קצב הנשימה והעומק נקבעים בעיקר על ידי לחץ חלקי עורקי של פחמן דו חמצני ולא על ידי העורק לחץ חלקי של חמצן, שמותר להשתנות בטווח רחב למדי לפני שמרכזי הנשימה ב- medulla oblongata ו- pons מגיבים אליו לשינוי קצב ועומק הנשימה.[6]
תרגיל מגביר את קצב הנשימה עקב תוספת הפחמן הדו-חמצני המיוצר על ידי חילוף החומרים המשופר של השרירים המתאמנים.[28] בנוסף תנועות פסיביות של הגפיים גם מייצרות בצורה רפלקסיבית עלייה בקצב הנשימה.[6][28]
מידע שהתקבל מ למתוח קולטנים בגבולות הריאות נפח גאות ושפל (עומק השאיפה והנשיפה).
תגובות ללחצים אטמוספריים נמוכים
ה alveoli פתוחים (דרך דרכי הנשימה) לאטמוספירה, וכתוצאה מכך לחץ אוויר מכתשי זהה לחלוטין ללחץ האוויר הסביבתי בגובה הים, בגובה או בכל אווירה מלאכותית (למשל תא צלילה, או תא לחץ דם) שהאדם נושם בחופשיות. עם התרחבות הריאות האוויר הלועבורי תופס נפח גדול יותר, שלו לחץ נופל באופן פרופורציונלי, מה שגורם לאוויר לזרום דרך דרכי הנשימה, עד שהלחץ בכבדות נמצא שוב בלחץ האוויר הסביבתי. ההפך קורה בזמן הנשיפה. זֶה תהליך (של שאיפה ונשיפה) זהה לחלוטין בגובה פני הים, כמו על גבי הר אוורסט, או ב תא צלילה אוֹ תא דקומפרסיה.
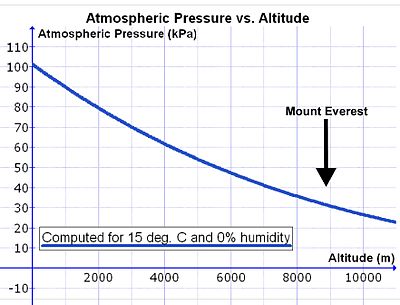
עם זאת, ככל שעולים מעל פני הים צפיפות האוויר פוחתת באופן אקספוננציאלי (ראה איור 14), במחציתם בערך עם כל 5500 מ 'גובה.[29] מכיוון שהרכב האוויר האטמוספרי כמעט קבוע מתחת ל 80 ק"מ, כתוצאה מהשפעת הערבוב המתמשכת של מזג האוויר, ריכוז החמצן באוויר (mmols O2 לליטר אוויר סביבתי) יורד באותו קצב כמו ירידת לחץ האוויר בגובה.[30] לכן, על מנת לנשום את אותה כמות חמצן לדקה, האדם צריך לשאוף נפח אוויר גדול יחסית לדקה בגובה מאשר בגובה פני הים. זה מושג על ידי נשימה עמוקה ומהירה יותר (כלומר היפרפנאה ) מאשר בגובה פני הים (ראה להלן).

עם זאת, יש סיבוך שמגדיל את נפח האוויר שצריך לשאוף לדקה (נפח דקה נשימתי ) לספק את אותה כמות חמצן לריאות בגובה כמו בגובה פני הים. במהלך שאיפה האוויר מחומם ורווי באדי מים במהלך מעברו דרך האזור מעברי אף ו לוֹעַ. לחץ אדי מים רווי תלוי רק בטמפרטורה. בטמפרטורת ליבת גוף של 37 מעלות צלזיוס זה 6.3kPa (47.0 מ"מ כספית), ללא קשר להשפעות אחרות, כולל גובה.[31] לפיכך, בגובה פני הים, כאשר לחץ האטמוספירה הסביבתי הוא כ- 100 kPa, האוויר הרטוב שזורם לריאות מה קנה הנשימה מורכב מאדי מים (6.3 kPa), חנקן (74.0 kPa), חמצן (19.7 kPa) וכמויות עקבות של פחמן דו חמצני וגזים אחרים (סה"כ 100 kPa). באוויר יבש את לחץ חלקי של O2 בגובה פני הים 21.0 kPa (כלומר 21% מ- 100 kPa), לעומת 19.7 kPa של חמצן שנכנס לאוויר המכתשי. (הלחץ החלקי בקנה הנשימה של חמצן הוא 21% מ- [100 kPa - 6.3 kPa] = 19.7 kPa). בפסגת הר אוורסט (בגובה 8,848 מ 'או 29,029 רגל) סך הכל לחץ האטמוספירה הוא 33.7 kPa, מתוכם 7.1 kPa (או 21%) הוא חמצן.[29] לאוויר הנכנס לריאות יש גם לחץ כולל של 33.7 kPa, מתוכם 6.3 kPa הם, באופן בלתי נמנע, אדי מים (כפי שהוא בגובה פני הים). זה מקטין את הלחץ החלקי של החמצן שנכנס לאלבולות ל -5.8 kPa (או 21% מ- [33.7 kPa - 6.3 kPa] = 5.8 kPa). הפחתת הלחץ החלקי של החמצן באוויר הנשאף גדולה אפוא באופן משמעותי מהמצביע על הפחתת הלחץ האטמוספרי הכולל בגובה (בהר אוורסט: 5.8 kPa לעומת. 7.1 kPa).
סיבוך מינורי נוסף קיים בגובה. אם נפח הריאות יוכפל באופן מיידי בתחילת השאיפה, לחץ האוויר בתוך הריאות היה חצוי. זה קורה בלי קשר לגובה. לפיכך, חציית לחץ האוויר בגובה פני הים (100 kPa) מביאה ללחץ אוויר תוך-ריאתי של 50 kPa. כשאתה עושה את אותו הדבר ב 5500 מ ', שם הלחץ האטמוספרי הוא 50 kPa בלבד, לחץ האוויר התוך-ריאי נופל ל -25 kPa. לכן, אותו שינוי בנפח הריאות בגובה פני הים גורם להפרש של 50 kPa בלחץ בין האוויר הסביבתי לאוויר התוך-ריאות, ואילו זה מביא להפרש של 25 kPa בלבד ב 5500 מ '. לחץ הנהיגה שמכריח אוויר לריאות בזמן שאיפה פוחת ולכן בגובה זה. ה ציון הזרמת האוויר לריאות במהלך שאיפה בגובה פני הים היא אפוא פי שניים מזו המתרחשת בגובה 5500 מ '. עם זאת, במציאות, שאיפה ונשיפה מתרחשות בצורה עדינה ופחות פתאומית בהרבה מאשר בדוגמה שניתנה. ההבדלים בין הלחצים האטמוספריים לתוך הריאה, המניעים אוויר פנימה ומחוצה להם מהריאות במהלך מחזור הנשימה, הם באזור של 2-3 kPa בלבד.[18][19] הכפלת או יותר מהבדלי הלחץ הקטנים הללו יכולה להיות מושגת רק על ידי שינויים גדולים מאוד במאמץ הנשימה בגבהים גדולים.
כל ההשפעות הנ"ל של לחצים נמוכים באטמוספירה על הנשימה מאופיינות בעיקר על ידי נשימה עמוקה ומהירה יותר (היפרפנאה ). המידה המדויקת של hyperpnea נקבעת על ידי הומאוסטט גז בדם, המסדיר את לחצים חלקיים של חמצן ופחמן דו חמצני בדם העורקי. זֶה הומאוסטט מעדיף את ויסות העורק לחץ חלקי של פחמן דו חמצני על פני חמצן בגובה פני הים.[6] כלומר, בגובה פני הים לחץ החלקי העורקי של CO2 נשמר על קרוב מאוד ל 5.3 kPa (או 40 מ"מ כספית) במגוון רחב של נסיבות, על חשבון הלחץ החלקי העורקי של O2, שמותר להשתנות בטווח רחב מאוד של ערכים, לפני שגורם לתגובת אוורור מתקנת. עם זאת, כאשר הלחץ האטמוספרי (ולכן הלחץ החלקי של O2 באוויר הסביבתי) נופל מתחת ל 50-75% מערכו בגובה פני הים, חמצן הומאוסטזיס מקבל עדיפות על פני הומאוסטזיס של פחמן דו חמצני.[6] מעבר זה מתרחש בגובה של כ 2500 מ '(או כ 8000 רגל). אם מתג זה מתרחש באופן פתאומי יחסית, hyperpnea בגובה רב יגרום לנפילה קשה בלחץ החלקי העורקי של פחמן דו חמצני, עם כתוצאה מעלייה ב- pH של פלזמה העורקים. זה תורם אחד ל- מחלה בגובה רב. מצד שני, אם המעבר להומאוסטזיס חמצן אינו שלם, אז היפוקסיה עשוי לסבך את התמונה הקלינית עם תוצאות שעלולות להיות קטלניות.
ישנם חיישני חמצן בקטן יותר סימפונות ו ברונכיולות. בתגובה ללחצים חלקיים נמוכים של חמצן באוויר הנשאף, חיישנים אלה גורמים להתכווצות עורקי הריאה באופן רפלקסיבי.[32] (זה בדיוק ההפך מהרפלקס המקביל ברקמות, שם לחץ חלקי עורקי נמוך של O2 לגרום להרחבת כלי דם עורקים.) בגובה זה גורם ל לחץ עורקי ריאתי לעלות וכתוצאה מכך התפלגות זרימת הדם לריאות הרבה יותר שווה ממה שקורה בגובה פני הים. בגובה פני הים לחץ העורקים הריאתי נמוך מאוד, והתוצאה היא ש צמרות הריאות מקבלים הרבה פחות דם מהבסיסים, שעוברות יתר על המידה של דם. רק באמצע הריאות זרימת דם ואוויר אל הכליות מותאמים באופן אידיאלי. בגובה וריאציה זו ב יחס אוורור / זלוף של alveoli מהחלק העליון של הריאות לתחתית מסולק, כאשר כל alveoli perfused ומאוורר בצורה פחות או יותר באופן האידיאלי מבחינה פיזיולוגית. זהו תורם חשוב נוסף ל- התאקלמות לגבהים גדולים and low oxygen pressures.
The kidneys measure the oxygen תוֹכֶן (mmol O2/liter blood, rather than the partial pressure of O2) of the arterial blood. When the oxygen content of the blood is chronically low, as at high altitude, the oxygen-sensitive kidney cells secrete אריתרופואטין (EPO) into the blood.[33][34] This hormone stimulates the red bone marrow to increase its rate of red cell production, which leads to an increase in the המטוקריט of the blood, and a consequent increase in its oxygen carrying capacity (due to the now high הֵמוֹגלוֹבִּין content of the blood). In other words, at the same arterial partial pressure of O2, a person with a high hematocrit carries more oxygen per liter of blood than a person with a lower hematocrit does. High altitude dwellers therefore have higher hematocrits than sea-level residents.[34][35]
Other functions of the lungs
Local defenses
Irritation of nerve endings within the מעברי האף אוֹ כיווני אוויר, can induce a cough reflex ו הִתעַטְשׁוּת. These responses cause air to be expelled forcefully from the קנה הנשימה אוֹ אף, בהתאמה. In this manner, irritants caught in the רִיר which lines the respiratory tract are expelled or moved to the פֶּה where they can be נִבלָע.[6] During coughing, contraction of the smooth muscle in the airway walls narrows the trachea by pulling the ends of the cartilage plates together and by pushing soft tissue into the lumen. This increases the expired airflow rate to dislodge and remove any irritant particle or mucus.
אפיתל נשימתי can secrete a variety of molecules that aid in the defense of the lungs. These include secretory אימונוגלובולינים (IgA), אספנים, defensins and other peptides and פרוטאזות, מיני חמצן תגובתי, ו מיני חנקן תגובתי. These secretions can act directly as antimicrobials to help keep the airway free of infection. מגוון רחב של כימוקינים ו ציטוקינים are also secreted that recruit the traditional immune cells and others to the site of infections.
חומר פעילי שטח immune function is primarily attributed to two proteins: SP-A and SP-D. These proteins can bind to sugars on the surface of pathogens and thereby opsonize them for uptake by phagocytes. It also regulates inflammatory responses and interacts with the adaptive immune response. Surfactant degradation or inactivation may contribute to enhanced susceptibility to lung inflammation and infection.[36]
Most of the respiratory system is lined with mucous membranes that contain רקמת לימפואיד הקשורה ברירית, שמייצר תאי דם לבנים כמו לימפוציטים.
Prevention of alveolar collapse
The lungs make a חומר פעילי שטח, a surface-active ליפופרוטאין complex (phospholipoprotein) formed by תאים מכתשיים מסוג II. It floats on the surface of the thin watery layer which lines the insides of the alveoli, reducing the water's surface tension.
The surface tension of a watery surface (the water-air interface) tends to make that surface shrink.[6] When that surface is curved as it is in the alveoli of the lungs, the shrinkage of the surface decreases the diameter of the alveoli. The more acute the curvature of the water-air interface the greater the tendency for the alveolus to collapse.[6] This has three effects. Firstly the surface tension inside the alveoli resists expansion of the alveoli during inhalation (i.e. it makes the lung stiff, or non-compliant). Surfactant reduces the surface tension and therefore makes the lungs more תואם, or less stiff, than if it were not there. Secondly, the diameters of the alveoli increase and decrease during the breathing cycle. This means that the alveoli have a greater tendency to collapse (i.e. cause אטלקטזיס ) at the end of exhalation that at the end of inhalation. Since surfactant floats on the watery surface, its molecules are more tightly packed together when the alveoli shrink during exhalation.[6] This causes them to have a greater surface tension-lowering effect when the alveoli are small than when they are large (as at the end of inhalation, when the surfactant molecules are more widely spaced). The tendency for the alveoli to collapse is therefore almost the same at the end of exhalation as at the end of inhalation. Thirdly, the surface tension of the curved watery layer lining the alveoli tends to draw water from the lung tissues into the alveoli. Surfactant reduces this danger to negligible levels, and keeps the alveoli dry.[6][37]
Pre-term babies who are unable to manufacture surfactant have lungs that tend to collapse each time they breathe out. Unless treated, this condition, called תסמונת מצוקה נשימתית, is fatal. Basic scientific experiments, carried out using cells from chicken lungs, support the potential for using סטֵרֵאוֹדִים as a means of furthering development of type II alveolar cells.[38] In fact, once a לידה מוקדמת is threatened, every effort is made to delay the birth, and a series of סטרואידים injections is frequently administered to the mother during this delay in an effort to promote lung maturation.[39]
Contributions to whole body functions
The lung vessels contain a fibrinolytic system that dissolves clots that may have arrived in the pulmonary circulation by תַסחִיף, often from the deep veins in the legs. They also release a variety of substances that enter the systemic arterial blood, and they remove other substances from the systemic venous blood that reach them via the pulmonary artery. כמה פרוסטגלנדינים are removed from the circulation, while others are synthesized in the lungs and released into the blood when lung tissue is stretched.
The lungs activate one hormone. The physiologically inactive decapeptide אנגיוטנסין אני is converted to the אלדוסטרון -releasing octapeptide, אנגיוטנסין II, in the pulmonary circulation. The reaction occurs in other tissues as well, but it is particularly prominent in the lungs. Angiotensin II also has a direct effect on arteriolar walls, causing arteriolar התכווצות כלי דם, and consequently a rise in לחץ דם עורקי.[40] Large amounts of the אנזים הממיר אנגיוטנסין responsible for this activation are located on the surfaces of the תאי אנדותל of the alveolar capillaries. The converting enzyme also inactivates ברדיקינין. Circulation time through the alveolar capillaries is less than one second, yet 70% of the angiotensin I reaching the lungs is converted to angiotensin II in a single trip through the capillaries. Four other peptidases have been identified on the surface of the pulmonary endothelial cells.
קוליות
The movement of gas through the גָרוֹן, לוֹעַ ו פֶּה allows humans to לְדַבֵּר, או phonate. Vocalization, or singing, in birds occurs via the סירינקס, an organ located at the base of the trachea. The vibration of air flowing across the larynx (מיתרי קול ), in humans, and the syrinx, in birds, results in sound. Because of this, gas movement is vital for תִקשׁוֹרֶת מטרות.
בקרת טמפרטורה
הִתנַשְׁמוּת in dogs, cats, birds and some other animals provides a means of reducing body temperature, by evaporating saliva in the mouth (instead of evaporating sweat on the skin).
משמעות קלינית
Disorders of the respiratory system can be classified into several general groups:
- Airway obstructive conditions (e.g., נַפַּחַת, בְּרוֹנכִיטִיס, אַסְתְמָה )
- Pulmonary restrictive conditions (e.g., לַיֶפֶת, סרקואידוזיס, alveolar damage, תפליט פלאורלי )
- Vascular diseases (e.g., בצקת ריאות, תסחיף ריאתי, יתר לחץ דם ריאתי )
- Infectious, environmental and other "diseases" (e.g., דלקת ריאות, שַׁחֶפֶת, אסבסטוזיס, particulate pollutants )
- Primary cancers (e.g. bronchial carcinoma, מזותליומה )
- Secondary cancers (e.g. cancers that originated elsewhere in the body, but have seeded themselves in the lungs)
- Insufficient surfactant (e.g. תסמונת מצוקה נשימתית in pre-term babies) .
Disorders of the respiratory system are usually treated by a ריאתי ו מטפל בנשימה.
Where there is an inability to breathe or an insufficiency in breathing a מאוורר רפואי עשוי להיות בשימוש.
Exceptional mammals
סוסים
Horses are obligate nasal breathers which means that they are different from many other mammals because they do not have the option of breathing through their mouths and must take in air through their noses.
פילים
ה פיל is the only mammal known to have no pleural space. במקום זאת, ה קָדקֳדִי ו צדר הרחם הקרביים are both composed of dense רקמת חיבור and joined to each other via loose connective tissue.[41] This lack of a pleural space, along with an unusually thick דִיאָפרַגמָה, are thought to be evolutionary adaptations allowing the elephant to remain underwater for long periods of time while breathing through its חדק which emerges as a snorkel.[42]
In the elephant the lungs are attached to the diaphragm and breathing relies mainly on the diaphragm rather than the expansion of the ribcage.[43]
ציפורים



מַפְתֵחַ:
1. גולגולת; 2. חוליות צוואר הרחם; 3. פורקולה; 4. קוראקואיד; 5. vertebral ribs; 6. sternum and its keel; 7. פִּיקַת הַבֶּרֶך; 8. שׁוֹרֶשׁ הָרֶגֶל; 9. ספרות; 10. שׁוּקָה (טיביוטרסוס ); 11. שׁוֹקִית (טיביוטרסוס ); 12. עֶצֶם הַיָרֵך; 13. איסקיום (innominate ); 14. חֵיק (innominate); 15. איליום (innominate); 16. חוליות הזנב; 17. פיגוסטיל; 18. synsacrum; 19. עֶצֶם הַשֶׁכֶם; 20. חוליות הגב; 21. עצם הזרוע; 22. גוֹמֶד; 23. רַדִיוּס; 24. קרפוס (קרפומטקרפוס ); 25. מטקרפוס (קרפומטקרפוס ); 26. ספרות; 27. אלולה

The respiratory system of birds differs significantly from that found in mammals. Firstly, they have rigid lungs which do not expand and contract during the breathing cycle. Instead an extensive system of נאדיות (Fig. 15) distributed throughout their bodies act as the bellows drawing environmental air into the sacs, and expelling the spent air after it has passed through the lungs (Fig. 18).[44] Birds also do not have דיאפרגמות אוֹ חללי pleural.
Bird lungs are smaller than those in mammals of comparable size, but the air sacs account for 15% of the total body volume, compared to the 7% devoted to the alveoli which act as the bellows in mammals.[45]
Inhalation and exhalation are brought about by alternately increasing and decreasing the volume of the entire thoraco-abdominal cavity (or צלם ) using both their abdominal and costal muscles.[46][47][48] During inhalation the muscles attached to the vertebral ribs (Fig. 17) contract angling them forwards and outwards. This pushes the sternal ribs, to which they are attached at almost right angles, downwards and forwards, taking the סטרנום (with its prominent שִׁדְרִית ) in the same direction (Fig. 17). This increases both the vertical and transverse diameters of thoracic portion of the trunk. The forward and downward movement of, particularly, the אֲחוֹרִי end of the sternum pulls the abdominal wall downwards, increasing the volume of that region of the trunk as well.[46] The increase in volume of the entire trunk cavity reduces the air pressure in all the thoraco-abdominal air sacs, causing them to fill with air as described below.
During exhalation the external oblique muscle which is attached to the sternum and vertebral ribs קדמית, and to the pelvis (pubis and ilium in Fig. 17) מאחור (forming part of the abdominal wall) reverses the inhalatory movement, while compressing the abdominal contents, thus increasing the pressure in all the air sacs. Air is therefore expelled from the respiratory system in the act of exhalation.[46]

During inhalation air enters the קנה הנשימה via the nostrils and mouth, and continues to just beyond the סירינקס at which point the trachea branches into two סימפונות ראשוניות, going to the two lungs (Fig. 16). The primary bronchi enter the lungs to become the intrapulmonary bronchi, which give off a set of parallel branches called ventrobronchi and, a little further on, an equivalent set of dorsobronchi (Fig. 16).[46] The ends of the intrapulmonary bronchi discharge air into the posterior air sacs at the זנב end of the bird. Each pair of dorso-ventrobronchi is connected by a large number of parallel microscopic air capillaries (or parabronchi ) איפה חילופי גז occurs (Fig. 16).[46] As the bird inhales, tracheal air flows through the intrapulmonary bronchi into the posterior air sacs, as well as into the דורסוbronchi, but not into the ventrobronchi (Fig. 18). This is due to the bronchial architecture which directs the inhaled air away from the openings of the ventrobronchi, into the continuation of the intrapulmonary bronchus towards the dorsobronchi and posterior air sacs.[50][51][52] From the dorsobronchi the inhaled air flows through the parabronchi (and therefore the gas exchanger) to the ventrobronchi from where the air can only escape into the expanding anterior air sacs. So, during inhalation, both the posterior and anterior air sacs expand,[46] the posterior air sacs filling with fresh inhaled air, while the anterior air sacs fill with "spent" (oxygen-poor) air that has just passed through the lungs.
During exhalation the pressure in the posterior air sacs (which were filled with fresh air during inhalation) increases due to the contraction of the oblique muscle described above. The aerodynamics of the interconnecting openings from the posterior air sacs to the dorsobronchi and intrapulmonary bronchi ensures that the air leaves these sacs in the direction of the lungs (via the dorsobronchi), rather than returning down the intrapulmonary bronchi (Fig. 18).[50][52] From the dorsobronchi the fresh air from the posterior air sacs flows through the parabronchi (in the same direction as occurred during inhalation) into ventrobronchi. The air passages connecting the ventrobronchi and anterior air sacs to the intrapulmonary bronchi direct the "spent", oxygen poor air from these two organs to the trachea from where it escapes to the exterior.[46] Oxygenated air therefore flows constantly (during the entire breathing cycle) in a single direction through the parabronchi.[53]
The blood flow through the bird lung is at right angles to the flow of air through the parabronchi, forming a cross-current flow exchange system (Fig. 19).[44][46][49] ה לחץ חלקי של חמצן in the parabronchi declines along their lengths as O2 diffuses into the blood. The blood capillaries leaving the exchanger near the entrance of airflow take up more O2 than do the capillaries leaving near the exit end of the parabronchi. When the contents of all capillaries mix, the final partial pressure of oxygen of the mixed pulmonary venous blood is higher than that of the exhaled air,[46][49] but is nevertheless less than half that of the inhaled air,[46] thus achieving roughly the same systemic arterial blood partial pressure of oxygen as mammals do with their bellows-type lungs.[46]
The trachea is an area of שטח מת: the oxygen-poor air it contains at the end of exhalation is the first air to re-enter the posterior air sacs and lungs. בהשוואה ל mammalian respiratory tract, the dead space volume in a bird is, on average, 4.5 times greater than it is in mammals of the same size.[45][46] Birds with long necks will inevitably have long tracheae, and must therefore take deeper breaths than mammals do to make allowances for their greater dead space volumes. In some birds (e.g. the ברבור, סיגנוס סיגנוס, ה white spoonbill, Platalea leucorodia, ה מנוף זנב, גראס אמריקנה, וה helmeted curassow, פויי פואסי) the trachea, which some cranes can be 1.5 m long,[46] is coiled back and forth within the body, drastically increasing the dead space ventilation.[46] The purpose of this extraordinary feature is unknown.
זוחלים

ה anatomical structure של ה ריאות is less complex in זוחלים מאשר ב יונקים, with reptiles lacking the very extensive airway tree structure found in mammalian lungs. חילופי גז in reptiles still occurs in alveoli למרות זאת.[44] Reptiles do not possess a דִיאָפרַגמָה. Thus, breathing occurs via a change in the volume of the body cavity which is controlled by contraction of שרירי צלעי in all reptiles except צבים. In turtles, contraction of specific pairs of flank muscles governs שְׁאִיפָה ו נְשִׁיפָה.[54]
דו-חיים
Both the lungs and the עור serve as respiratory organs in דו-חיים. The ventilation of the lungs in amphibians relies on אוורור לחץ חיובי. Muscles lower the floor of the oral cavity, enlarging it and drawing in air through the nostrils into the חלל פה. With the nostrils and mouth closed, the floor of the oral cavity is then pushed up, which forces air down the trachea into the lungs. The skin of these animals is highly vascularized and moist, with moisture maintained via secretion of רִיר from specialised cells, and is involved in נשימה עורית. While the lungs are of primary organs for gas exchange between the blood and the environmental air (when out of the water), the skin's unique properties aid rapid gas exchange when amphibians are submerged in oxygen-rich water.[55]Some amphibians have gills, either in the early stages of their development (e.g. ראשנים שֶׁל צפרדעים ), while others retain them into adulthood (e.g. some סלמנדרות ).[44]
דג


Oxygen is poorly soluble in water. Fully aerated מים מתוקים therefore contains only 8–10 ml O2/liter compared to the O2 concentration of 210 ml/liter in the air at sea level.[59] יתר על כן, coefficient of diffusion (i.e. the rate at which a substances diffuses from a region of high concentration to one of low concentration, under standard conditions) of the respiratory gases is typically 10,000 faster in air than in water.[59] Thus oxygen, for instance, has a diffusion coefficient of 17.6 mm2/s in air, but only 0.0021 mm2/s in water.[60][61][62][63] The corresponding values for carbon dioxide are 16 mm2/s in air and 0.0016 mm2/s in water.[62][63] This means that when oxygen is taken up from the water in contact with a gas exchanger, it is replaced considerably more slowly by the oxygen from the oxygen-rich regions small distances away from the exchanger than would have occurred in air. Fish have developed זימים deal with these problems. Gills are specialized organs containing חוטים, which further divide into למלות. The lamellae contain a dense thin walled capillary network that exposes a large gas exchange surface area to the very large volumes of water passing over them.[64]
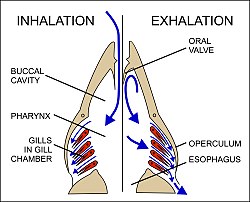
Gills use a החלפת זרם נגד system that increases the efficiency of oxygen-uptake from the water.[56][57][58] Fresh oxygenated water taken in through the mouth is uninterruptedly "pumped" through the gills in one direction, while the blood in the lamellae flows in the opposite direction, creating the countercurrent blood and water flow (Fig. 22), on which the fish's survival depends.[58]
Water is drawn in through the mouth by closing the אופרקולום (gill cover), and enlarging the mouth cavity (Fig. 23). Simultaneously the gill chambers enlarge, producing a lower pressure there than in the mouth causing water to flow over the gills.[58] The mouth cavity then contracts inducing the closure of the passive oral valves, thereby preventing the back-flow of water from the mouth (Fig. 23).[58][65] The water in the mouth is, instead, forced over the gills, while the gill chambers contract emptying the water they contain through the opercular openings (Fig. 23). Back-flow into the gill chamber during the inhalatory phase is prevented by a membrane along the ventroposterior border of the operculum (diagram on the left in Fig. 23). Thus the mouth cavity and gill chambers act alternately as suction pump and pressure pump to maintain a steady flow of water over the gills in one direction.[58] Since the blood in the lamellar capillaries flows in the opposite direction to that of the water, the consequent countercurrent flow of blood and water maintains steep concentration gradients for oxygen and carbon dioxide along the entire length of each capillary (lower diagram in Fig. 22). Oxygen is, therefore, able to continually diffuse down its gradient into the blood, and the carbon dioxide down its gradient into the water.[57] Although countercurrent exchange systems theoretically allow an almost complete transfer of a respiratory gas from one side of the exchanger to the other, in fish less than 80% of the oxygen in the water flowing over the gills is generally transferred to the blood.[56]
In certain active פלגי sharks, water passes through the mouth and over the gills while they are moving, in a process known as "ram ventilation".[66] While at rest, most sharks pump water over their gills, as most bony fish do, to ensure that oxygenated water continues to flow over their gills. But a small number of species have lost the ability to pump water through their gills and must swim without rest. מינים אלה הם חייבים מאווררי איל ויהיה ככל הנראה לחנוק אם אינך מסוגל לזוז. אוורור איל חובה נכון גם לגבי מינים דגים גרמיים פלגניים.[67]
There are a few fish that can obtain oxygen for brief periods of time from air swallowed from above the surface of the water. לכן דגי ריאות possess one or two lungs, and the labyrinth fish have developed a special "labyrinth organ", which characterizes this suborder of fish. The labyrinth organ is a much-folded supraענפי אבזר breathing organ. It is formed by a כלי דם expansion of the epibranchial bone of the first gill arch, and is used for נשימה באוויר.[68]
This organ allows labyrinth fish to take in חַמצָן directly from the air, instead of taking it from the water in which they reside through use of זימים. The labyrinth organ helps the oxygen in the inhaled air to be absorbed into the זרם הדם. As a result, labyrinth fish can survive for a short period of time out of water, as they can inhale the air around them, provided they stay moist.
Labyrinth fish are not born with functional labyrinth organs. The development of the organ is gradual and most juvenile labyrinth fish breathe entirely with their gills and develop the labyrinth organs when they grow older.[68]
חסרי חוליות
פרוקי רגליים
מינים מסוימים של סרטן use a respiratory organ called a branchiostegal lung.[69] Its gill-like structure increases the surface area for gas exchange which is more suited to taking oxygen from the air than from water. Some of the smallest עכבישים ו קרדית can breathe simply by exchanging gas through the surface of the body. Larger spiders, עקרבים ואחר פרוקי רגליים use a primitive book lung.
חרקים
Most insects breath passively through their ספירקלות (special openings in the שלד חיצוני ) and the air reaches every part of the body by means of a series of smaller and smaller tubes called 'trachaea' when their diameters are relatively large, and 'קנה הנשימה ' when their diameters are very small. The tracheoles make contact with individual cells throughout the body.[44] They are partially filled with fluid, which can be withdrawn from the individual tracheoles when the tissues, such as muscles, are active and have a high demand for oxygen, bringing the air closer to the active cells.[44] This is probably brought about by the buildup of lactic acid in the active muscles causing an osmotic gradient, moving the water out of the tracheoles and into the active cells. Diffusion of gases is effective over small distances but not over larger ones, this is one of the reasons insects are all relatively small. Insects which do not have spiracles and trachaea, such as some Collembola, breathe directly through their skins, also by diffusion of gases.[70]
The number of spiracles an insect has is variable between species, however, they always come in pairs, one on each side of the body, and usually one pair per segment. Some of the Diplura have eleven, with four pairs on the thorax, but in most of the ancient forms of insects, such as Dragonflies and Grasshoppers there are two thoracic and eight abdominal spiracles. However, in most of the remaining insects, there are fewer. It is at the level of the tracheoles that oxygen is delivered to the cells for respiration.
Insects were once believed to exchange gases with the environment continuously by the simple diffusion of gases into the tracheal system. More recently, however, large variation in insect ventilatory patterns has been documented and insect respiration appears to be highly variable. Some small insects do not demonstrate continuous respiratory movements and may lack muscular control of the spiracles. Others, however, utilize כיווץ שרירים של ה בֶּטֶן along with coordinated spiracle contraction and relaxation to generate cyclical gas exchange patterns and to reduce water loss into the atmosphere. The most extreme form of these patterns is termed discontinuous gas exchange מחזורים.[71]
רכיכות
רכיכות generally possess gills that allow gas exchange between the aqueous environment and their circulatory systems. These animals also possess a heart that pumps blood containing המוציאנין as its oxygen-capturing molecule.[44] Hence, this respiratory system is similar to that of vertebrate fish. ה respiratory system of gastropods can include either gills or a lung.
צמחים
צמחים להשתמש פחמן דו חמצני gas in the process of פוטוסינתזה, and exhale חַמצָן gas as waste. The chemical equation of photosynthesis is 6 CO2 (carbon dioxide) and 6 H2O (water), which in the presence of sunlight makes C6ה12או6 (glucose) and 6 O2 (חַמצָן). Photosynthesis uses electrons on the carbon atoms as the repository for the energy obtained from sunlight.[72] Respiration is the opposite of photosynthesis. It reclaims the energy to power chemical reactions in cells. In so doing the carbon atoms and their electrons are combined with oxygen forming CO2 which is easily removed from both the cells and the organism. Plants use both processes, photosynthesis to capture the energy and מטבוליזם חמצוני to use it.
Plant respiration is limited by the process of ריכוך. Plants take in carbon dioxide through holes, known as סטומטה, that can open and close on the undersides of their משאיר and sometimes other parts of their anatomy. Most plants require some oxygen for קטבולית processes (break-down reactions that release energy). But the quantity of O2 used per hour is small as they are not involved in activities that require high rates of אירובי חילוף חומרים. Their requirement for air, however, is very high as they need CO2 for photosynthesis, which constitutes only 0.04% of the environmental air. Thus, to make 1 g of glucose requires the removal of all the CO2 מ לפחות 18.7 liters of air at sea level. But inefficiencies in the photosynthetic process cause considerably greater volumes of air to be used.[72][73]
ראה גם
- אירוע חמצון נהדר – Paleoproterozoic surge in atmospheric oxygen
- הסתגלות לנשימה
- ספירומטריה
- בדיקת תפקוד ריאתי (PFT)
הפניות
- ^ Campbell, Neil A. (1990). ביולוגיה (מהדורה שנייה). Redwood City, Calif.: Benjamin/Cummings Pub. Co. pp. 834–835. ISBN 0-8053-1800-3.
- ^ Hsia, CC; Hyde, DM; Weibel, ER (15 במרץ 2016). "Lung Structure and the Intrinsic Challenges of Gas Exchange". פיזיולוגיה מקיפה. 6 (2): 827–95. דוי:10.1002/cphy.c150028. PMC 5026132. PMID 27065169.
- ^ West, John B. (1995). Respiratory physiology-- the essentials. בולטימור: וויליאמס ווילקינס. עמ.1–10. ISBN 0-683-08937-4.
- ^ א ב Gilroy, Anne M.; MacPherson, Brian R.; Ross, Lawrence M. (2008). Atlas of Anatomy. שטוטגרט: תימה. עמ '108–111. ISBN 978-1-60406-062-1.
- ^ א ב Pocock, Gillian; Richards, Christopher D. (2006). פיזיולוגיה אנושית: בסיס הרפואה (מהדורה שלישית). אוקספורד: הוצאת אוניברסיטת אוקספורד. עמ '315–317. ISBN 978-0-19-856878-0.
- ^ א ב ג ד ה f ז ח אני j k l M נ o עמ ' ש ר ס t u v טורטורה, ג'רארד ג'יי; Anagnostakos, Nicholas P. (1987). עקרונות האנטומיה והפיזיולוגיה (מהדורה חמישית). New York: Harper & Row, Publishers. עמ.556–586. ISBN 0-06-350729-3.
- ^ קצמרק, רוברט מ.; Dimas, Steven; Mack, Craig W. (13 August 2013). Essentials of Respiratory Care - E-Book. מדעי הבריאות אלסבייה. ISBN 9780323277785.
- ^ Netter, Frank H. (2014). אטלס האנטומיה האנושית כולל ייעוץ והנחיות אינטראקטיביות לסטודנטים (מהדורה 6). פילדלפיה, פן: W B Saunders Co. עמ ' 200. ISBN 978-1-4557-0418-7.
- ^ מטון, אנתאה; ז'אן הופקינס; צ'רלס וויליאם מקלופלין; סוזן ג'ונסון; מריאנה קווון וורנר; דייויד להארט; ג'יל ד 'רייט (1993). ביולוגיה אנושית ובריאות. wood Cliffs, New Jersey, USA: Prentice Hall. ISBN 0-13-981176-1.[דף נדרש ]
- ^ א ב ג Fowler W.S. (1948). "Lung Function studies. II. The respiratory dead space". אמ. ג'י פיסיול. 154 (3): 405–416. דוי:10.1152/ajplegacy.1948.154.3.405. PMID 18101134.
- ^ "anatomical dead space". TheFreeDictionary.com.
- ^ א ב טורטורה, ג'רארד ג'יי; Anagnostakos, Nicholas P. (1987). עקרונות האנטומיה והפיזיולוגיה (מהדורה חמישית). New York: Harper & Row, Publishers. עמ.570–572. ISBN 0-06-350729-3.
- ^ טורובסקי, ג'ייסון (29/04/2016). "האם אתה צריך לנשום דרך הפה או האף שלך?". קליבלנד קליניק. הוחזר 2020-06-28.
- ^ "Your Nose, the Guardian of Your Lungs". המרכז הרפואי בוסטון. הוחזר 2020-06-29.
- ^ Dahl, Melissa (2011-01-11). "'Mouth-breathing' gross, harmful to your health". חדשות NBC. הוחזר 2020-06-28.
- ^ Valcheva, Zornitsa (January 2018). "THE ROLE OF MOUTH BREATHING ON DENTITION DEVELOPMENT AND FORMATION" (PDF). Journal of IMAB. הוחזר 2020-05-31.
- ^ Gross, Terry (2020-05-27). "How The 'Lost Art' Of Breathing Can Impact Sleep And Resilience". הרדיו הציבורי הלאומי (NPR) /אוויר צח. הוחזר 2020-06-23.
- ^ א ב Koen, Chrisvan L.; Koeslag, Johan H. (1995). "On the stability of subatmospheric intrapleural and intracranial pressures". חדשות במדעי הפיזיולוגיה. 10 (4): 176–178. דוי:10.1152/physiologyonline.1995.10.4.176.
- ^ א ב West, J.B. (1985). Respiratory physiology: the essentials. בולטימור: וויליאמס ווילקינס. pp. 21–30, 84–84, 98–101.
- ^ Burke, TV; Küng, M; Burki, NK (1989). "Pulmonary gas exchange during histamine-induced bronchoconstriction in asthmatic subjects". חזה. 96 (4): 752–6. דוי:10.1378/chest.96.4.752. PMID 2791669. S2CID 18569280.
- ^ Taylor, D (1996). "The Valsalva Manoeuvre: A critical review". דרום האגודה לרפואה תת-מימית באוקיינוס השקט. 26 (1). ISSN 0813-1988. OCLC 16986801. הוחזר 14 במרץ 2016.
- ^ מטון, אנתאה; הופקינס, ז'אן סוזן; ג'ונסון, צ'רלס וויליאם; מקלפלין, מריאנה קווון; וורנר, דייוויד; להארט רייט, ג'יל (2010). ביולוגיה אנושית ובריאות. צוקי אנגלווד: אולם פרנטיס. עמ '108–118. ISBN 978-0134234359.
- ^ א ב ג Williams, Peter L.; Warwick, Roger; Dyson, Mary; Bannister, Lawrence H. (1989). האנטומיה של גריי (מהדורה שלושים ושבע). אדינבורו: צ'רצ'יל ליווינגסטון. pp. 1278–1282. ISBN 0443-041776.
- ^ Lovelock, James (1991). Healing Gaia: Practical medicine for the Planet. ניו יורק: הרמוניה ספרים. pp. 21–34, 73–88. ISBN 0-517-57848-4.
- ^ Shu, BC; Chang, YY; Lee, FY; Tzeng, DS; Lin, HY; Lung, FW (2007-10-31). "Parental attachment, premorbid personality, and mental health in young males with hyperventilation syndrome". מחקר פסיכיאטריה. 153 (2): 163–70. דוי:10.1016/j.psychres.2006.05.006. PMID 17659783. S2CID 3931401.
- ^ Henry RP, Swenson ER (June 2000). "The distribution and physiological significance of carbonic anhydrase in vertebrate gas exchange organs". פיזיולוגיה של הנשימה. 121 (1): 1–12. דוי:10.1016/S0034-5687(00)00110-9. PMID 10854618.
- ^ Diem, K.; Lentner, C. (1970). "דם - חומרים אנאורגניים". ב: טבלאות מדעיות (מהדורה שביעית). Basle, Switzerland: CIBA-GEIGY Ltd. p. 571.
- ^ א ב "Respiration". Harvey Project. הוחזר 27 ביולי 2012.
- ^ א ב "Online high altitude oxygen calculator". altitude.org. הועבר לארכיון מ המקורי ב- 29 ביולי 2012. הוחזר 15 באוגוסט 2007.
- ^ Tyson, P.D.; Preston-White, R.A. (2013). The weather and climate of Southern Africa. קייפטאון: הוצאת אוניברסיטת אוקספורד. pp. 3–10, 14–16, 360. ISBN 9780195718065.
- ^ Diem, K.; Lenter, C. (1970). Scientific Tables (מהדורה שביעית). Basle, Switzerland: Ciba-Geigy. עמ '257–258.
- ^ Von Euler, U.S.; Liljestrand, G. (1946). "Observations on the pulmonary arterial blood pressure in the cat". Acta Physiologica Scandinavica. 12 (4): 301–320. דוי:10.1111/j.1748-1716.1946.tb00389.x.
- ^ "EPO Detection". הסוכנות העולמית למניעת סמים. הוחזר 7 בספטמבר 2017.
- ^ א ב טורטורה, ג'רארד ג'יי; Anagnostakos, Nicholas P. (1987). עקרונות האנטומיה והפיזיולוגיה (מהדורה חמישית). New York: Harper & Row, Publishers. עמ.444–445. ISBN 0-06-350729-3.
- ^ Fisher JW, Koury S, Ducey T, Mendel S (1996). "Erythropoietin production by interstitial cells of hypoxic monkey kidneys". כתב העת הבריטי להמטולוגיה. 95 (1): 27–32. דוי:10.1046/j.1365-2141.1996.d01-1864.x. PMID 8857934. S2CID 38309595.
- ^ Wright, Jo Rae (2004). "Host Defense Functions of Pulmonary Surfactant". ביולוגיה של הילוד. 85 (4): 326–32. דוי:10.1159/000078172. PMID 15211087. S2CID 25469141.
- ^ West, John B. (1994). Respiratory physiology-- the essentials. בולטימור: וויליאמס ווילקינס. עמ.21–30, 84–84, 98–101. ISBN 0-683-08937-4.
- ^ Sullivan, LC; Orgeig, S (2001). "Dexamethasone and epinephrine stimulate surfactant secretion in type II cells of embryonic chickens". כתב העת האמריקני לפיזיולוגיה. פיזיולוגיה רגולטורית, אינטגרטיבית והשוואתית. 281 (3): R770–7. דוי:10.1152/ajpregu.2001.281.3.r770. PMID 11506991.
- ^ Premature Babies, Lung Development & Respiratory Distress Syndrome. Pregnancy-facts.com.
- ^ Kanaide, Hideo; Ichiki, Toshihiro; Nishimura, Junji; Hirano, Katsuya (2003-11-28). "Cellular Mechanism of Vasoconstriction Induced by Angiotensin II It Remains To Be Determined". מחקר מחזור. 93 (11): 1015–1017. דוי:10.1161/01.RES.0000105920.33926.60. ISSN 0009-7330. PMID 14645130.
- ^ West, John B.; Ravichandran (1993). "Snorkel breathing in the elephant explains the unique anatomy of its pleura". פיזיולוגיה של הנשימה. 126 (1): 1–8. דוי:10.1016/S0034-5687(01)00203-1. PMID 11311306.
- ^ ווסט, ג'ון ב '(2002). "Why doesn't the elephant have a pleural space?". News Physiol Sci. 17 (2): 47–50. דוי:10.1152 / nips.01374.2001. PMID 11909991. S2CID 27321751.
- ^ Shoshani, Jeheskel (December 1998). "Understanding proboscidean evolution: a formidable task". מגמות באקולוגיה ואבולוציה. 13 (12): 480–487. דוי:10.1016/S0169-5347(98)01491-8. PMID 21238404.
- ^ א ב ג ד ה f ז Campbell, Neil A. (1990). ביולוגיה (מהדורה שנייה). Redwood City, Calif.: Benjamin/Cummings Pub. Co. pp. 836–844. ISBN 0-8053-1800-3.
- ^ א ב Whittow, G. Causey (2000). Sturkie's Avian Physiology. סן דייגו, קליפורניה: עיתונות אקדמית. pp. 233–241. ISBN 978-0-12-747605-6.
- ^ א ב ג ד ה f ז ח אני j k l M נ o ריצ'סון, ג'י "BIO 554/754 - אורניתולוגיה: נשימה בעופות". המחלקה למדעים ביולוגיים, אוניברסיטת קנטקי המזרחית. הוחזר 2009-04-23.
- ^ Storer, Tracy I.; Usinger, R. L.; סטיבינס, רוברט סי .; Nybakken, James W. (1997). General Zoology (מהדורה שישית). ניו יורק: מקגרו-היל. עמ.752–753. ISBN 0-07-061780-5.
- ^ Romer, Alfred Sherwood (1970). The Vertebrate body (מהדורה רביעית). פילדלפיה: W.B. סונדרס. עמ.323–324. ISBN 0-7216-7667-7.
- ^ א ב ג סקוט, גרהאם ר '(2011). "פרשנות: ביצועים גבוהים: הפיזיולוגיה הייחודית של ציפורים שעפות בגובה רב". כתב העת לביולוגיה ניסיונית. 214 (Pt 15): 2455–2462. דוי:10.1242 / jeb.052548. PMID 21753038.
- ^ א ב מיינה, ג'ון נ '(2005). מערכת שק האוויר הריאה של התפתחות ציפורים, מבנה ותפקודם; עם 6 שולחנות. ברלין: ספרינגר. עמ '3.2–3.3 "ריאה", "מערכת דרכי הנשימה (ברונכיול)" 66–82. ISBN 978-3-540-25595-6.
- ^ Krautwald-Junghanns, Maria-Elisabeth; ואח '. (2010). Diagnostic Imaging of Exotic Pets: Birds, Small Mammals, Reptiles. Germany: Manson Publishing. ISBN 978-3-89993-049-8.
- ^ א ב Sturkie, P.D. (1976). Sturkie, P. D (ed.). Avian Physiology. ניו יורק: ספרינגר ורלאג. עמ ' 201. דוי:10.1007/978-1-4612-4862-0. ISBN 978-1-4612-9335-4. S2CID 36415426.
- ^ Ritchison, Gary. "Ornithology (Bio 554/754):Bird Respiratory System". אוניברסיטת קנטקי המזרחית. אוחזר 27-06-2007.
- ^ מערכת נשימה. אנציקלופדיה בריטניקה.
- ^ Gottlieb, G; Jackson DC (1976). "Importance of pulmonary ventilation in respiratory control in the bullfrog". Am J Physiol. 230 (3): 608–13. דוי:10.1152/ajplegacy.1976.230.3.608. PMID 4976.
- ^ א ב ג Campbell, Neil A. (1990). ביולוגיה (מהדורה שנייה).רדווד סיטי, קליפורניה: חברת פרסום בנימין / קאמינגס, בע"מ עמ '836–838. ISBN 0-8053-1800-3.
- ^ א ב ג יוז GM (1972). "מורפומטריה של זימי דגים". פיזיולוגיה של הנשימה. 14 (1–2): 1–25. דוי:10.1016 / 0034-5687 (72) 90014-x. PMID 5042155.
- ^ א ב ג ד ה f סטורר, טרייסי הראשון .; Usinger, R. L.; סטיבינס, רוברט סי .; Nybakken, James W. (1997). זואולוגיה כללית (מהדורה שישית). ניו יורק: מקגרו-היל. עמ.668–670. ISBN 0-07-061780-5.
- ^ א ב מ. ב. נ 'רוברטס; מייקל רייס; גרייס מונגר (2000). ביולוגיה מתקדמת. לונדון, בריטניה: נלסון. עמ '164–165.
- ^ Cussler, E. L. (1997). דיפוזיה: העברה המונית במערכות נוזלים (מהדורה שנייה). ניו יורק: הוצאת אוניברסיטת קיימברידג '. ISBN 0-521-45078-0.
- ^ וולטי, ג'יימס ר .; וויקס, צ'רלס א. וילסון, רוברט א. רורר, גרגורי (2001). יסודות המומנטום, החום והעברת המונים. וויילי. ISBN 978-0-470-12868-8.
- ^ א ב CRC Press Online: מדריך CRC לכימיה ופיזיקה, סעיף 6, מהדורה 91
- ^ א ב ריכוך
- ^ ניוסטד ג'יימס די (1967). "מבנה עדין של למות הנשימה של זימים טלאוסטאניים". חקר תאים ורקמות. 79 (3): 396–428. דוי:10.1007 / bf00335484. PMID 5598734. S2CID 20771899.
- ^ רומר, אלפרד שרווד; פרסונס, תומאס ס '(1977). גוף החוליות. פילדלפיה, הרשות הפלסטינית: הולט-סונדרס אינטרנשיונל. עמ '316–327. ISBN 0-03-910284-X.
- ^ גילברטסון, לאנס (1999). מדריך מעבדה לזואולוגיה. ניו יורק: מקגרו-היל. ISBN 0-07-237716-X.
- ^ וויליאם ג'יי בנטה (1996). "נשימה עמוקה". הוחזר 2007-08-28.
- ^ א ב פינטר, ה '(1986). דג מבוך. הסדרה החינוכית של בארון בע"מ, ISBN 0-8120-5635-3
- ^ הלפרין J, אנסאלדו M, פלרנו GN, Luquet CM (יולי 2000). "נשימה דו-מודאלית בסרטן האסטרה Chasmagnathus granulatus Dana 1851 - מחקרים פיזיולוגיים ומורפולוגיים". ביוכימיה ופיזיולוגיה השוואתית. חלק א ', פיזיולוגיה מולקולרית ואינטגרטיבית. 126 (3): 341–9. דוי:10.1016 / S1095-6433 (00) 00216-6. PMID 10964029.
- ^ רשת החיים של כדור הארץ, מורפולוגיה חרקים ואנטומיה. Earthlife.net. אוחזר בתאריך 21/04/2013.
- ^ לייטון, JRB (ינואר 1996). "החלפת גז לא רציפה בחרקים". Annu Rev Entomol. 41: 309–324. דוי:10.1146 / annurev.en.41.010196.001521. PMID 8546448.
- ^ א ב מייבש שיער, לוברט (1995). "פוטוסינתזה". בתוך: ביוכימיה (מהדורה רביעית). ניו יורק: W.H. FreeMan and Company. עמ '653-680. ISBN 0-7167-2009-4.
- ^ קמפבל, ניל א '(1990). ביולוגיה (מהדורה שנייה). רדווד סיטי, קליפורניה: חברת פרסום בנימין / קאמינגס, בע"מ עמ '206-223. ISBN 0-8053-1800-3.
קישורים חיצוניים
- תיאור ברמת התיכון של מערכת הנשימה
- מבוא למערכת הנשימה
- עזרה מדעית: מערכת הנשימה מדריך פשוט לתלמידי תיכון
- מערכת הנשימה רמת אוניברסיטה (מסמך Microsoft Word)
- הרצאות בפיזיולוגיה של הנשימה על ידי פיזיולוג נשימה ידוע ג'ון ב. ווסט (גם ב יוטיוב )
| משאבי הספרייה על אודות מערכת נשימה |
| בקרת רשות |
|---|